

The molecular identities of the Caenorhabditis elegans intraflagellar transport genes dyf-6, daf-10 and osm-1
- PMID: 16648645
- PMCID: PMC1526656
- DOI: 10.1534/genetics.106.056721
The molecular identities of the Caenorhabditis elegans intraflagellar transport genes dyf-6, daf-10 and osm-1
Abstract
The Caenorhabditis elegans genes dyf-6, daf-10, and osm-1 are among the set of genes that affect chemotaxis and the ability of certain sensory neurons to take up fluorescent dyes from the environment. Some genes in this category are known to be required for intraflagellar transport (IFT), which is the bidirectional movement of raft-like particles along the axonemes of cilia and flagella. The cloning of dyf-6, daf-10, and osm-1 are described here. The daf-10 and osm-1 gene products resemble each other and contain WD and WAA repeats. DYF-6, the product of a complex locus, lacks known motifs, but orthologs are present in flies and mammals. Phenotypic analysis of dyf-6 mutants expressing an OSM-6::GFP reporter indicates that the cilia of the amphid and phasmid dendritic endings are foreshortened. Consistent with genetic mosaic analysis, which indicates that dyf-6 functions in neurons of the amphid sensilla, DYF-6::GFP is expressed in amphid and phasmid neurons. Movement of DYF-6::GFP within the ciliated endings of the neurons indicates that DYF-6 is involved in IFT. In addition, IFT can be observed in dauer larvae.
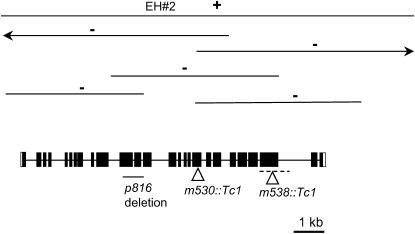
Figures








References
-
- Albert, P. S., and D. L. Riddle, 1983. Developmental alterations in sensory neuroanatomy of the Caenorhabditis elegans dauer larva. J. Comp. Neurol. 219: 461–481. - PubMed
-
- Albert, P. S., S. J. Brown and D. L. Riddle, 1981. Sensory control of dauer larva formation in Caenorhabditis elegans. J. Comp. Neurol. 198: 435–451. - PubMed
-
- Avidor-Reiss, T., A. M. Maer, E. Koundakjian, A. Polyanovsky, T. Keil et al., 2004. Decoding cilia function: defining specialized genes required for compartmentalized cilia biogenesis. Cell 117: 527–539. - PubMed
-
- Bargmann, C. I., and H. R. Horvitz, 1991. Control of larval development by chemosensory neurons in Caenorhabditis elegans. Science 251: 1243–1246. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
