

A mitotic topoisomerase II checkpoint in budding yeast is required for genome stability but acts independently of Pds1/securin
- PMID: 16651657
- PMCID: PMC1472475
- DOI: 10.1101/gad.1367206
A mitotic topoisomerase II checkpoint in budding yeast is required for genome stability but acts independently of Pds1/securin
Abstract
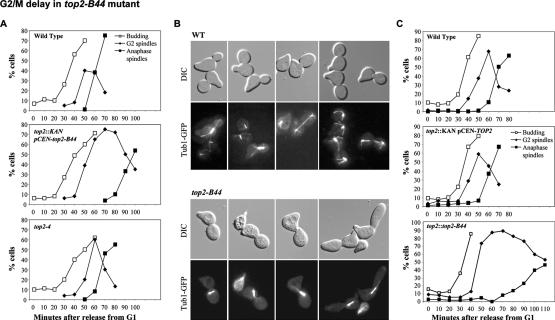
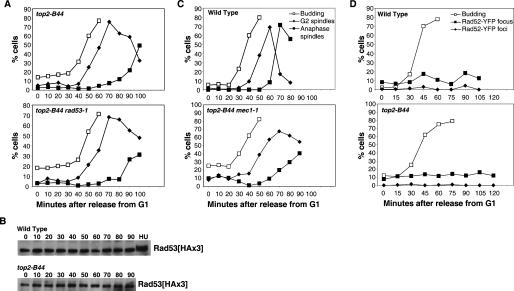
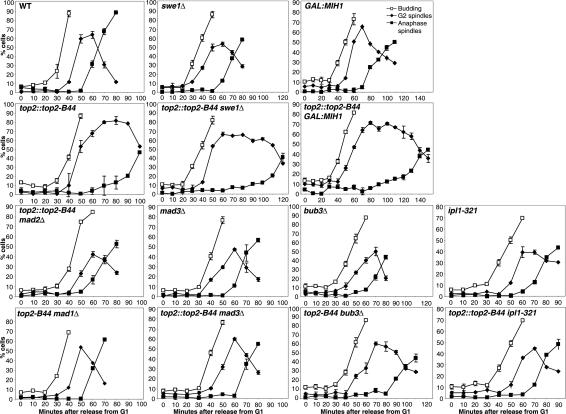
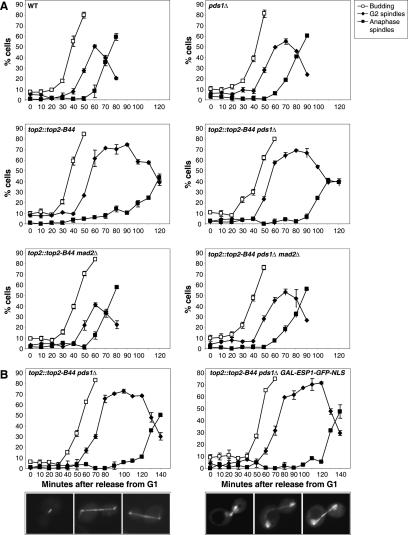
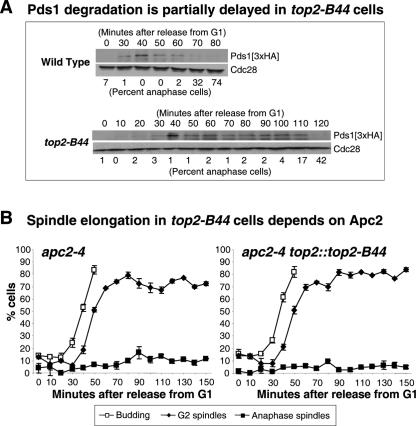
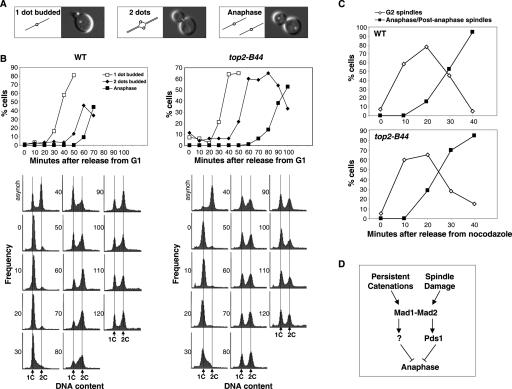
Topoisomerase II (Topo II) performs topological modifications on double-stranded DNA molecules that are essential for chromosome condensation, resolution, and segregation. In mammals, G2 and metaphase cell cycle delays induced by Topo II poisons have been proposed to be the result of checkpoint activation in response to the catenation state of DNA. However, the apparent lack of such controls in model organisms has excluded genetic proof that Topo II checkpoints exist and are separable from the conventional DNA damage checkpoint controls. But here, we define a Topo II-dependent G2/M checkpoint in a genetically amenable eukaryote, budding yeast, and demonstrate that this checkpoint enhances cell survival. Conversely, a lack of the checkpoint results in aneuploidy. Neither DNA damage-responsive pathways nor Pds1/securin are needed for this checkpoint. Unusually, spindle assembly checkpoint components are required for the Topo II checkpoint, but checkpoint activation is not the result of failed chromosome biorientation or a lack of spindle tension. Thus, compromised Topo II function activates a yeast checkpoint system that operates by a novel mechanism.
Figures
References
-
- Adachi Y., Luke M., Laemmli U.K. Chromosome assembly in vitro: Topoisomerase II is required for condensation. Cell. 1991;64:137–148. - PubMed
-
- Adachi N., Iiizumi S., So S., Koyama H. Genetic evidence for involvement of two distinct nonhomologous end-joining pathways in repair of topoisomerase II-mediated DNA damage. Biochem. Biophys. Res. Commun. 2004;318:856–861. - PubMed
-
- Bachant J., Alcasabas A., Blat Y., Kleckner N., Elledge S.J. The SUMO-1 isopeptidase Smt4 is linked to centromeric cohesion through SUMO-1 modification of DNA topoisomerase II. Mol. Cell. 2002;9:1169–1182. - PubMed
-
- Charron M., Hancock R. DNA topoisomerase II is required for formation of mitotic chromosomes in Chinese hamster ovary cells: Studies using the inhibitor 4′-demethylepipodophyllotoxin 9-(4,6-O-thenylidene-β-D-glucopyranoside). Biochemistry. 1990;29:9531–9537. - PubMed
-
- Clarke D.J., Giménez-Abián J.F. Checkpoints controlling mitosis. Bioessays. 2000;22:351–363. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases