

SOX2 is a dose-dependent regulator of retinal neural progenitor competence
- PMID: 16651659
- PMCID: PMC1472477
- DOI: 10.1101/gad.1407906
SOX2 is a dose-dependent regulator of retinal neural progenitor competence
Abstract
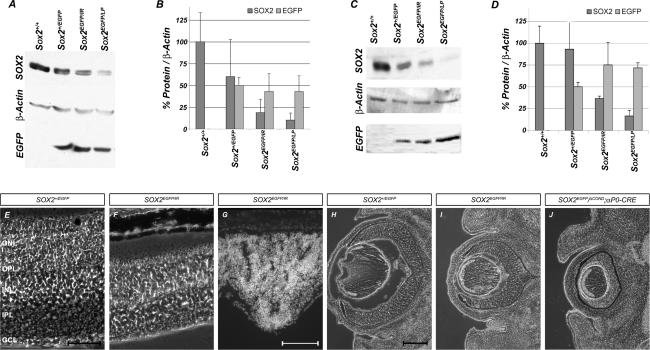
Approximately 10% of humans with anophthalmia (absent eye) or severe microphthalmia (small eye) show haploid insufficiency due to mutations in SOX2, a SOXB1-HMG box transcription factor. However, at present, the molecular or cellular mechanisms responsible for these conditions are poorly understood. Here, we directly assessed the requirement for SOX2 during eye development by generating a gene-dosage allelic series of Sox2 mutations in the mouse. The Sox2 mutant mice display a range of eye phenotypes consistent with human syndromes and the severity of these phenotypes directly relates to the levels of SOX2 expression found in progenitor cells of the neural retina. Retinal progenitor cells with conditionally ablated Sox2 lose competence to both proliferate and terminally differentiate. In contrast, in Sox2 hypomorphic/null mice, a reduction of SOX2 expression to <40% of normal causes variable microphthalmia as a result of aberrant neural progenitor differentiation. Furthermore, we provide genetic and molecular evidence that SOX2 activity, in a concentration-dependent manner, plays a key role in the regulation of the NOTCH1 signaling pathway in retinal progenitor cells. Collectively, these results show that precise regulation of SOX2 dosage is critical for temporal and spatial regulation of retinal progenitor cell differentiation and provide a cellular and molecular model for understanding how hypomorphic levels of SOX2 cause retinal defects in humans.
Figures







References
-
- Ahmad I., Dooley C.M., Polk D.L. Delta-1 is a regulator of neurogenesis in the vertebrate retina. Dev. Biol. 1997;185:92–103. - PubMed
-
- Austin C.P., Feldman D.E., Ida J.A., Jr., Cepko C.L. Vertebrate retinal ganglion cells are selected from competent progenitors by the action of Notch. Development. 1995;121:3637–3650. - PubMed
-
- Bylund M., Andersson E., Novitch B.G., Muhr J. Vertebrate neurogenesis is counteracted by Sox1-3 activity. Nat. Neurosci. 2003;6:1162–1168. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases