

RNA interference analysis of Legionella in Drosophila cells: exploitation of early secretory apparatus dynamics
- PMID: 16652170
- PMCID: PMC1447669
- DOI: 10.1371/journal.ppat.0020034
RNA interference analysis of Legionella in Drosophila cells: exploitation of early secretory apparatus dynamics
Abstract
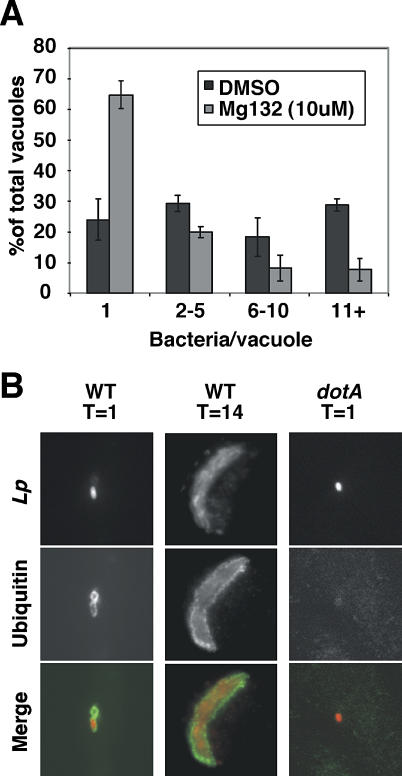
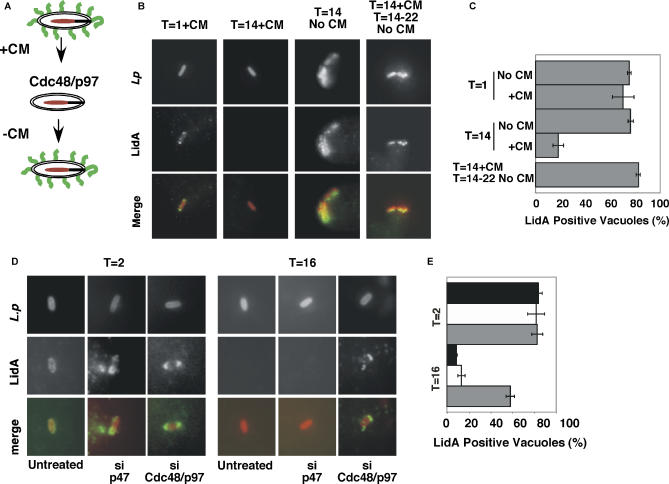
Legionella pneumophila translocates multiple bacterial effector proteins into host cells to direct formation of a replication vacuole for the bacterium. The emerging consensus is that formation of this compartment involves recruitment of membrane material that traffics between the endoplasmic reticulum (ER) and Golgi. To investigate this model, a targeted approach was used to knock down expression of proteins involved in membrane trafficking, using RNA interference in Drosophila cells. Surprisingly, few single knockdowns of ER-Golgi transport proteins decreased L. pneumophila replication. By analyzing double-stranded RNAs in pairs, combinations were identified that together caused defects in intracellular replication, consistent with the model that membrane traffic funnels into the replication vacuole from multiple sources. In particular, simultaneous depletion of the intermediate compartment and Golgi-tethering factor transport protein particle together with the ER SNARE protein Sec22 reduced replication efficiency, indicating that introduction of lesions at distinct sites in the secretory system reduces replication efficiency. In contrast to knockdowns in secretory traffic, which required multiple simultaneous hits, knockdown of single cytosolic components of ER-associated degradation, including Cdc48/p97 and associated cofactors, was sufficient to inhibit intracellular replication. The requirement for the Cdc48/p97 complex was conserved in mammalian cells, in which replication vacuoles showed intense recruitment of ubiquitinated proteins, the preferred substrates of Cdc48/p97. This complex promoted dislocation of both ubiquitinated proteins and bacterial effectors from the replication vacuole, consistent with the model that maintenance of high-level replication requires surveillance of the vacuole surface. This work demonstrates that L. pneumophila has the ability to gain access to multiple sites in the secretory system and provides the first evidence for a role of the Cdc48/p97 complex in promoting intracellular replication of pathogens and maintenance of replication vacuoles.
Conflict of interest statement
Figures





References
-
- Segal G, Shuman HA. Intracellular multiplication and human macrophage killing by Legionella pneumophila are inhibited by conjugal components of IncQ plasmid RSF1010. Mol Microbiol. 1998;30:197–208. - PubMed
-
- Vogel JP, Andrews HL, Wong SK, Isberg RR. Conjugative transfer by the virulence system of Legionella pneumophila . Science. 1998;279:873–876. - PubMed
-
- Nagai H, Kagan JC, Zhu X, Kahn RA, Roy CR. A bacterial guanine nucleotide exchange factor activates ARF on Legionella phagosomes. Science. 2002;295:679–682. - PubMed
-
- Conover GM, Derre I, Vogel JP, Isberg RR. The Legionella pneumophila LidA protein: A translocated substrate of the Dot/Icm system associated with maintenance of bacterial integrity. Molecular Microbiology. 2003;48:305–321. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
