

A phosphatidylinositol-3-kinase-dependent signal transition regulates ARF1 and ARF6 during Fcgamma receptor-mediated phagocytosis
- PMID: 16669702
- PMCID: PMC1457017
- DOI: 10.1371/journal.pbio.0040162
A phosphatidylinositol-3-kinase-dependent signal transition regulates ARF1 and ARF6 during Fcgamma receptor-mediated phagocytosis
Abstract
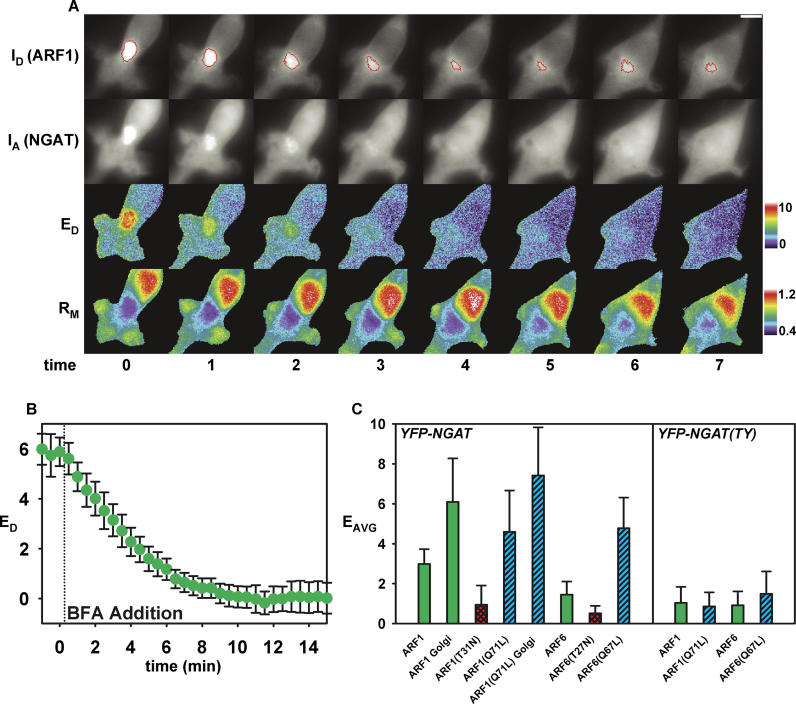
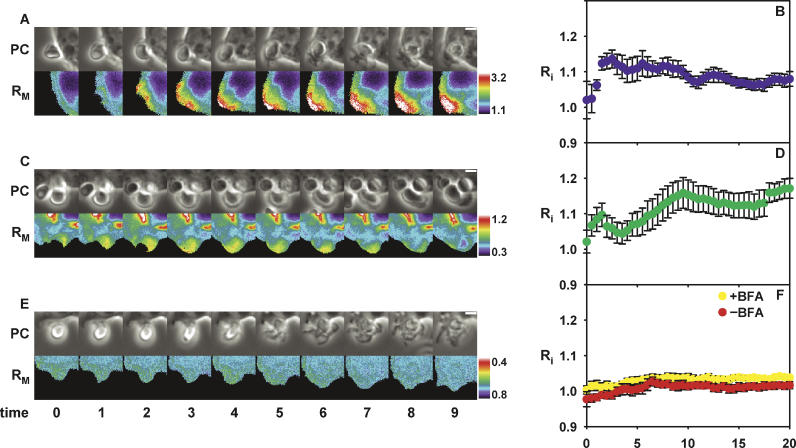
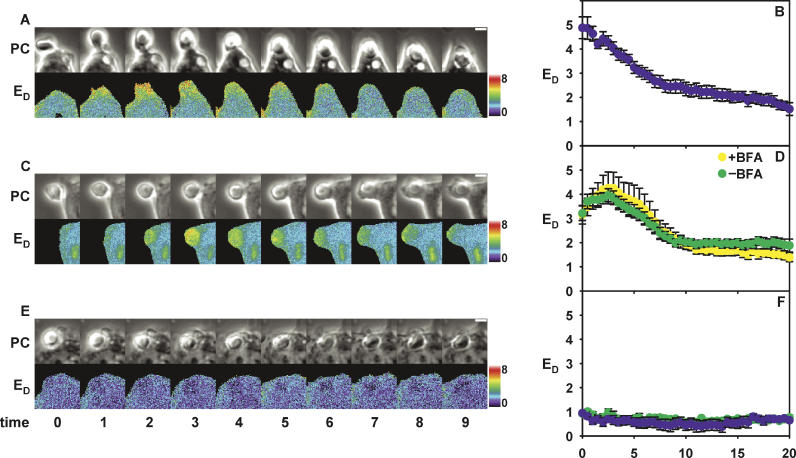
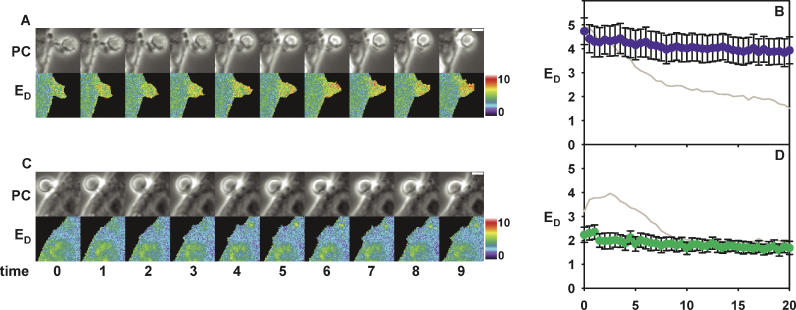
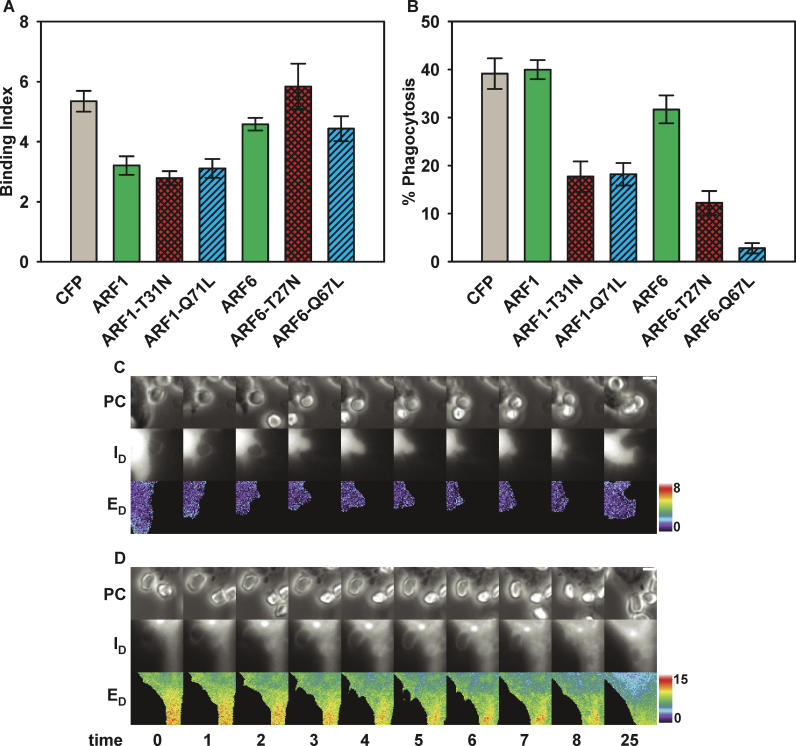
Fcgamma receptor (FcgammaR)-mediated phagocytosis of IgG-coated particles is regulated by 3'-phosphoinositides (3'PIs) and several classes of small GTPases, including ARF6 from the ADP Ribosylation Factor subfamily. The insensitivity of phagocytosis to brefeldin A (BFA), an inhibitor of certain ARF guanine nucleotide exchange factors (GEFs), previously indicated that ARF1 did not participate in phagocytosis. In this study, we show that ARF1 was activated during FcgammaR-mediated phagocytosis and that blocking normal ARF1 cycling inhibited phagosome closure. We examined the distributions and activation patterns of ARF6 and ARF1 during FcgammaR-mediated phagocytosis using fluorescence resonance energy transfer (FRET) stoichiometric microscopy of macrophages expressing CFP- or YFP-chimeras of ARF1, ARF6, and a GTP-ARF-binding protein domain. Both GTPases were activated by BFA-insensitive factors at sites of phagocytosis. ARF6 activation was restricted to the leading edge of the phagocytic cup, while ARF1 activation was delayed and delocalized over the phagosome. Phagocytic cups formed after inhibition of PI 3-kinase (PI-3K) contained persistently activated ARF6 and minimally activated ARF1. This indicates that a PI-3K-dependent signal transition defines the sequence of ARF GTPase activation during phagocytosis and that ARF6 and ARF1 coordinate different functions at the forming phagosome.
Figures
Comment in
-
Inconspicuous consumption: uncovering the molecular pathways behind phagocytosis.PLoS Biol. 2006 Jun;4(6):e190. doi: 10.1371/journal.pbio.0040190. Epub 2006 May 9. PLoS Biol. 2006. PMID: 20076587 Free PMC article. No abstract available.
References
-
- Cox D, Tseng CC, Bjekic G, Greenberg S. A requirement for phosphatidylinositol 3-kinase in pseudopod extension. J Biol Chem. 1999;274:1240–1247. - PubMed
-
- Zhang Q, Cox D, Tseng CC, Donaldson JG, Greenberg S. A requirement for ARF6 in Fcγ receptor–mediated phagocytosis in macrophages. J Biol Chem. 1998;273:19977–19981. - PubMed
-
- Honda A, Nogami M, Yokozeki T, Yamazaki M, Nakamura H, et al. Phosphatidylinositol 4-phosphate 5-kinase α is a downstream effector of the small G protein ARF6 in membrane ruffle formation. Cell. 1999;99:521–532. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
