

Mannan-binding lectin activates C3 and the alternative complement pathway without involvement of C2
- PMID: 16670774
- PMCID: PMC1451204
- DOI: 10.1172/JCI25982
Mannan-binding lectin activates C3 and the alternative complement pathway without involvement of C2
Abstract
Lectin pathway activation of C3 is known to involve target recognition by mannan-binding lectin (MBL) or ficolins and generation of classical pathway C3 convertase via cleavage of C4 and C2 by MBL-associated serine protease 2 (MASP-2). We investigated C3 activation in C2-deficient human sera and in sera with other defined defects of complement to assess other mechanisms through which MBL might recruit complement. The capacity of serum to support C3 deposition was examined by ELISA using microtiter plates coated with O antigen-specific oligosaccharides derived from Salmonella typhimurium, S. thompson, and S. enteritidis corresponding to serogroups B, C, and D (BO, CO, and DO). MBL bound to CO, but not to BO and DO, and efficiently supported C3 deposition in the absence of C2, C4, or MASP-2. The existence of an MBL-dependent C2 bypass mechanism for alternative pathway-mediated C3 activation was clearly demonstrated using CO, solid-phase mannan, and E. coli LPS. MASP-1 might contribute, but was not required for C3 deposition in the model used. Independent of MBL, specific antibodies to CO supported C3 deposition through classical and alternative pathways. MBL-dependent C2 bypass activation could be particularly important in various inherited and acquired complement deficiency states.
Figures
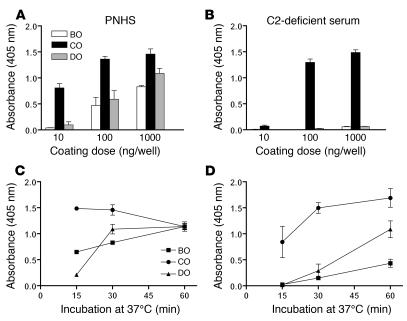
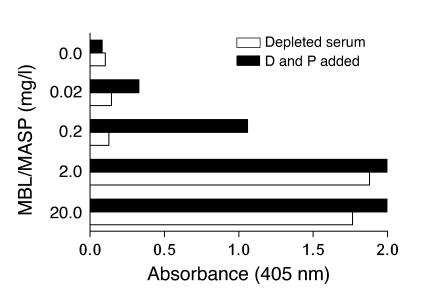
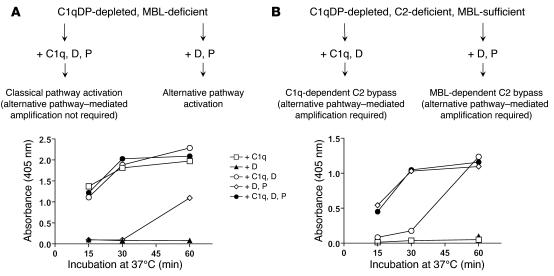
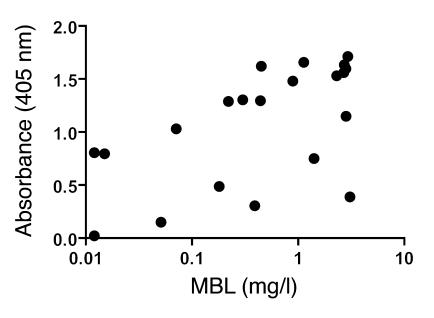
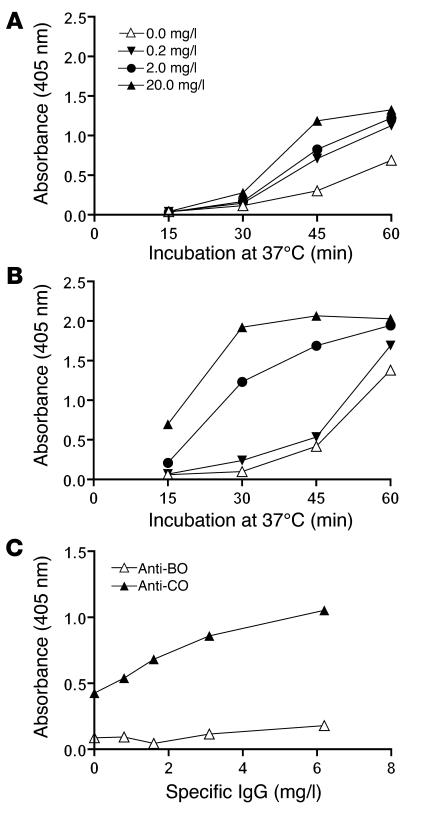
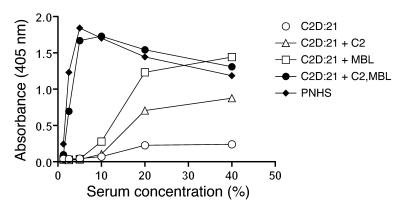
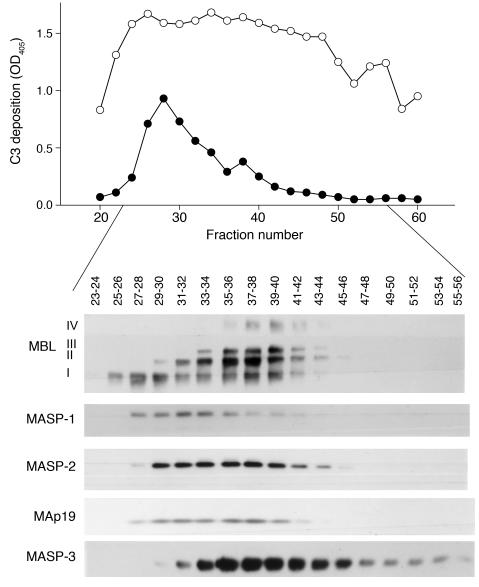
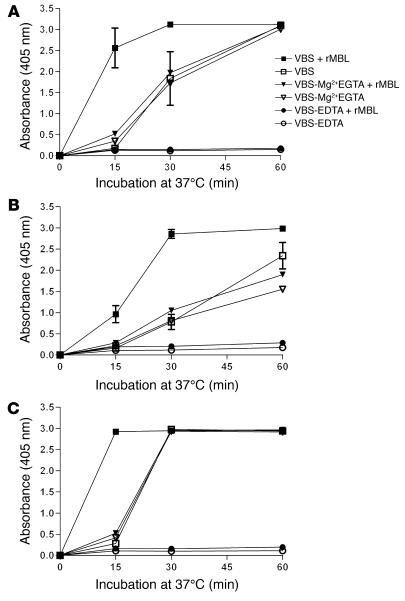
Comment in
- J Clin Invest. 116:1215.
References
-
- Turner M.W., Hamvas R.M. Mannose-binding lectin: structure, function, genetics and disease associations. Rev. Immunogenet. . 2000;39:305–322. - PubMed
-
- Hoffmann J.A., Kafator F.C., Janeway C.A., Ezekowitz R. A.B. Phylogenetic perspectives in innate immunity. Science. 1999;284:1313–1318. - PubMed
-
- Holmskov U., Thiel S., Jensenius J.C. Collectins and ficolins: humoral lectins of the innate immune defense. Annu. Rev. Immunol. 2003;21:547–578. - PubMed
-
- Thiel S., et al. A second serine protease associated with mannan-binding lectin that activates complement. Nature. . 1997;386:506–510. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
