

Multiple groESL operons are not key targets of RpoH1 and RpoH2 in Sinorhizobium meliloti
- PMID: 16672605
- PMCID: PMC1482865
- DOI: 10.1128/JB.188.10.3507-3515.2006
Multiple groESL operons are not key targets of RpoH1 and RpoH2 in Sinorhizobium meliloti
Abstract
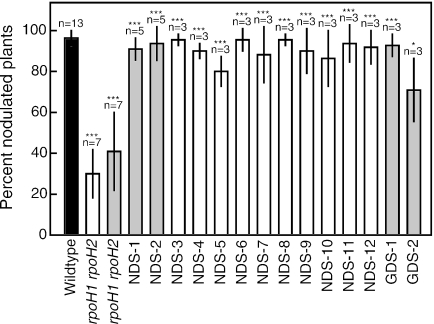
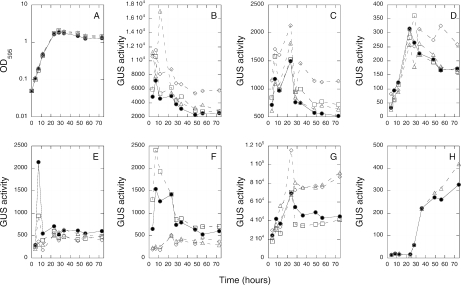
Among the rhizobia that establish nitrogen-fixing nodules on the roots of host plants, many contain multiple copies of genes encoding the sigma factor RpoH and the chaperone GroEL/GroES. In Sinorhizobium meliloti there are two rpoH genes, four groESL operons, and one groEL gene. rpoH1 mutants are defective for growth at high temperature and form ineffective nodules, rpoH1 rpoH2 double mutants are unable to form nodules, and groESL1 mutants form ineffective nodules. To explore the roles of RpoH1 and RpoH2, we identified mutants that suppress both the growth and nodulation defects. These mutants do not suppress the nitrogen fixation defect. This implies that the functions of RpoH1 during growth and RpoH1/RpoH2 during the initiation of symbiosis are similar but that there is a different function of RpoH1 needed later during symbiosis. We showed that, unlike in Escherichia coli, overexpression of groESL is not sufficient to bypass any of the RpoH defects. Under free-living conditions, we determined that RpoH2 does not control expression of the groE genes, and RpoH1 only controls expression of groESL5. Finally, we completed the series of groE mutants by constructing groESL3 and groEL4 mutants and demonstrated that they do not display symbiotic defects. Therefore, the only groESL operon required by itself for symbiosis is groESL1. Taken together, these results suggest that GroEL/GroES production alone cannot explain the requirements for RpoH1 and RpoH2 in S. meliloti and that there must be other crucial targets.
Figures


Similar articles
-
Dual RpoH sigma factors and transcriptional plasticity in a symbiotic bacterium.J Bacteriol. 2012 Sep;194(18):4983-94. doi: 10.1128/JB.00449-12. Epub 2012 Jul 6. J Bacteriol. 2012. PMID: 22773790 Free PMC article.
-
Only one of five groEL genes is required for viability and successful symbiosis in Sinorhizobium meliloti.J Bacteriol. 2007 Mar;189(5):1884-9. doi: 10.1128/JB.01542-06. Epub 2006 Dec 8. J Bacteriol. 2007. PMID: 17158666 Free PMC article.
-
Two RpoH homologs responsible for the expression of heat shock protein genes in Sinorhizobium meliloti.Mol Gen Genet. 2001 Feb;264(6):902-12. doi: 10.1007/s004380000380. Mol Gen Genet. 2001. PMID: 11254138
-
The Escherichia coli groE chaperonins.Semin Cell Biol. 1990 Feb;1(1):19-25. Semin Cell Biol. 1990. PMID: 1983267 Review.
-
Regulation and organization of the groE and dnaK operons in Eubacteria.FEMS Microbiol Lett. 1996 Apr 15;138(1):1-10. doi: 10.1111/j.1574-6968.1996.tb08126.x. FEMS Microbiol Lett. 1996. PMID: 8674965 Review.
Cited by
-
Dual RpoH sigma factors and transcriptional plasticity in a symbiotic bacterium.J Bacteriol. 2012 Sep;194(18):4983-94. doi: 10.1128/JB.00449-12. Epub 2012 Jul 6. J Bacteriol. 2012. PMID: 22773790 Free PMC article.
-
Role of specific quorum-sensing signals in the regulation of exopolysaccharide II production within Sinorhizobium meliloti spreading colonies.PLoS One. 2012;7(8):e42611. doi: 10.1371/journal.pone.0042611. Epub 2012 Aug 13. PLoS One. 2012. PMID: 22912712 Free PMC article.
-
Expression of Two RpoH Sigma Factors in Sinorhizobium meliloti upon Heat Shock.Microbes Environ. 2017 Dec 27;32(4):394-397. doi: 10.1264/jsme2.ME17087. Epub 2017 Dec 2. Microbes Environ. 2017. PMID: 29199214 Free PMC article.
-
Sinorhizobium meliloti sigma factors RpoE1 and RpoE4 are activated in stationary phase in response to sulfite.PLoS One. 2012;7(11):e50768. doi: 10.1371/journal.pone.0050768. Epub 2012 Nov 30. PLoS One. 2012. PMID: 23226379 Free PMC article.
-
The role of sigma factor RpoH1 in the pH stress response of Sinorhizobium meliloti.BMC Microbiol. 2010 Oct 18;10:265. doi: 10.1186/1471-2180-10-265. BMC Microbiol. 2010. PMID: 20955556 Free PMC article.
References
-
- Babst, M., H. Hennecke, and H.-M. Fischer. 1996. Two different mechanisms are involved in the heat-shock regulation of chaperonin gene expression in Bradyrhizobium japonicum. Mol. Microbiol. 19:827-839. - PubMed
-
- Barnett, M. J., V. Oke, and S. R. Long. 2000. New genetic tools for use in the Rhizobiaceae and other bacteria. BioTechniques 29:240-245. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
