

Endothelial nitric oxide synthase regulates microvascular hyperpermeability in vivo
- PMID: 16675496
- PMCID: PMC1817804
- DOI: 10.1113/jphysiol.2006.108175
Endothelial nitric oxide synthase regulates microvascular hyperpermeability in vivo
Abstract
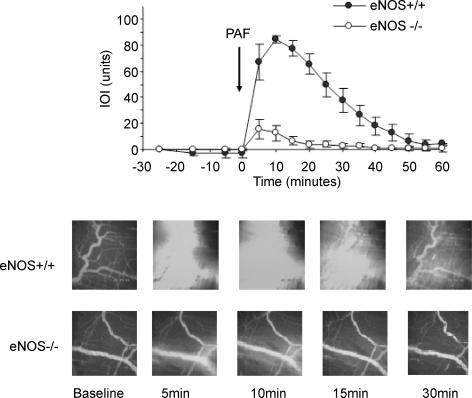
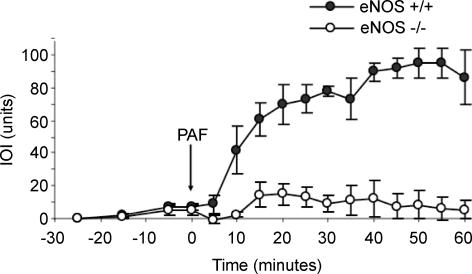
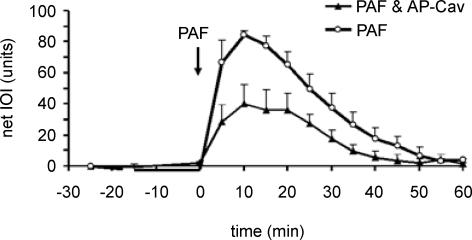
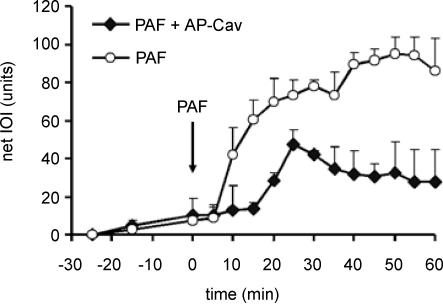
Nitric oxide (NO) is an important regulator of blood flow, but its role in permeability is still challenged. We tested in vivo the hypotheses that: (a) endothelial nitric oxide synthase (eNOS) is not essential for regulation of baseline permeability; (b) eNOS is essential for hyperpermeability responses in inflammation; and (c) molecular inhibition of eNOS with caveolin-1 scaffolding domain (AP-Cav) reduces eNOS-regulated hyperpermeability. We used eNOS-deficient (eNOS-/-) mice and their wild-type control as experimental animals, platelet-activating factor (PAF) at 10(-7) m as the test pro-inflammatory agent, and integrated optical intensity (IOI) as an index of microvascular permeability. PAF increased permeability in wild-type cremaster muscle from a baseline of 2.4 +/- 2.2 to a peak net value of 84.4 +/- 2.7 units, while the corresponding values in cremaster muscle of eNOS-/- mice were 1.0 +/- 0.3 and 15.6 +/- 7.7 units (P < 0.05). Similarly, PAF increased IOI in the mesentery of wild-type mice but much less in the mesentery of eNOS-/- mice. PAF increased IOI to comparable values in the mesenteries of wild-type mice and those lacking the gene for inducible NOS (iNOS). Administration of AP-Cav blocked the microvascular hyperpermeability responses to 10(-7) m PAF. We conclude that: (1) baseline permeability does not depend on eNOS; (2) eNOS and NO are integral elements of the signalling pathway for the hyperpermeability response to PAF; (3) iNOS does not affect either baseline permeability or hyperpermeability responses to PAF; and (4) caveolin-1 inhibits eNOS regulation of microvascular permeability in vivo. Our results establish eNOS as an important regulator of microvascular permeability in inflammation.
Figures
References
-
- Aramoto H, Breslin JW, Pappas PJ, Hobson RW, 2nd, Durán WN. Vascular endothelial growth factor stimulates differential signaling pathways in in vivo microcirculation. Am J Physiol Heart Circ Physiol. 2004;287:H1590–H1598. - PubMed
-
- Bekker AY, Ritter AB, Durán WN. Analysis of microvascular permeability to macromolecules by video-image digital processing. Microvasc Res. 1989;38:200–216. - PubMed
-
- Boric MP, Roblero JS, Durán WN. Quantitation of bradykinin-induced microvascular leakage of FITC-dextran in rat cremaster muscle. Microvasc Res. 1987;33:397–412. - PubMed
-
- Breslin JW, Pappas PJ, Cerveira JJ, Hobson RW, 2nd, Durán WN. VEGF increases endothelial permeability by separate signaling pathways involving ERK-1/2 and nitric oxide. Am J Physiol Heart Circ Physiol. 2003;284:H92–H100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical