

Androgens, progestins, and glucocorticoids induce follicle-stimulating hormone beta-subunit gene expression at the level of the gonadotrope
- PMID: 16675544
- PMCID: PMC2941439
- DOI: 10.1210/me.2005-0316
Androgens, progestins, and glucocorticoids induce follicle-stimulating hormone beta-subunit gene expression at the level of the gonadotrope
Abstract
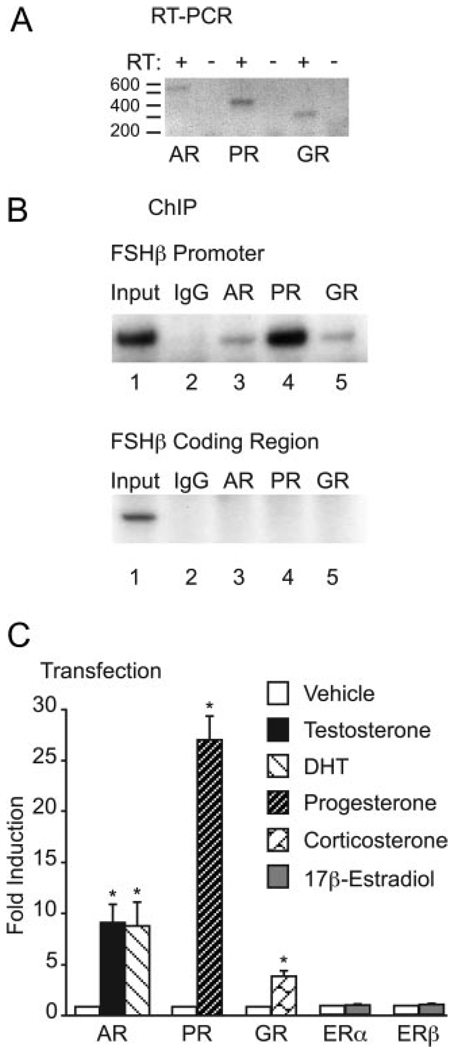
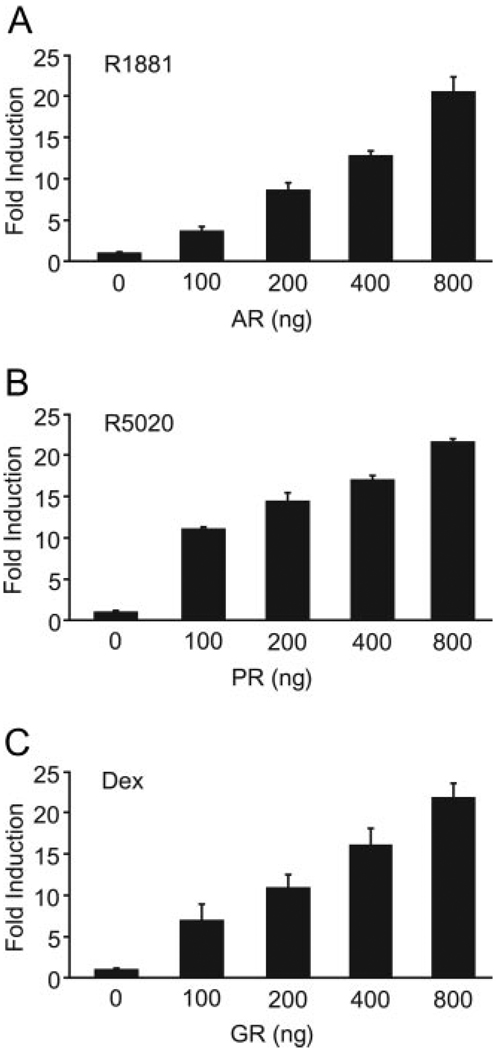
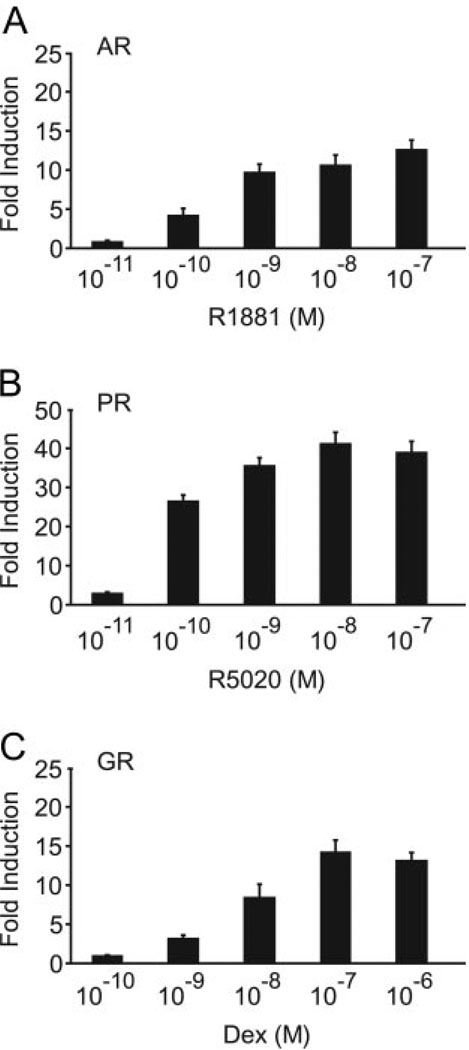
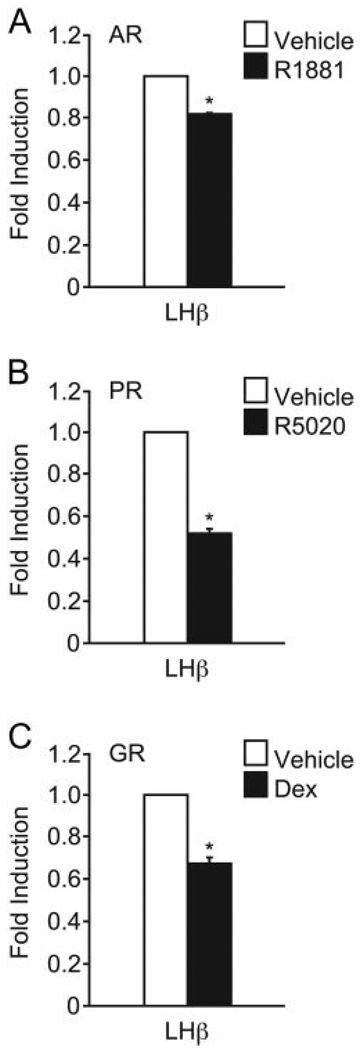
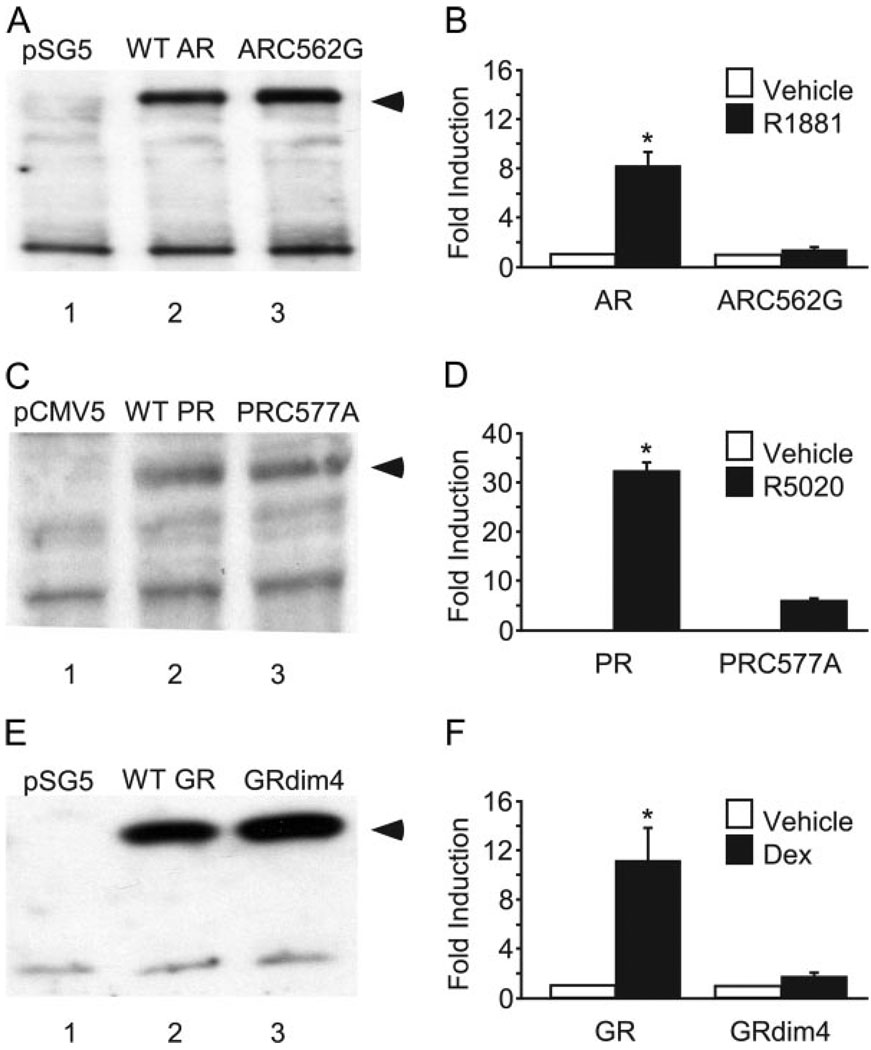
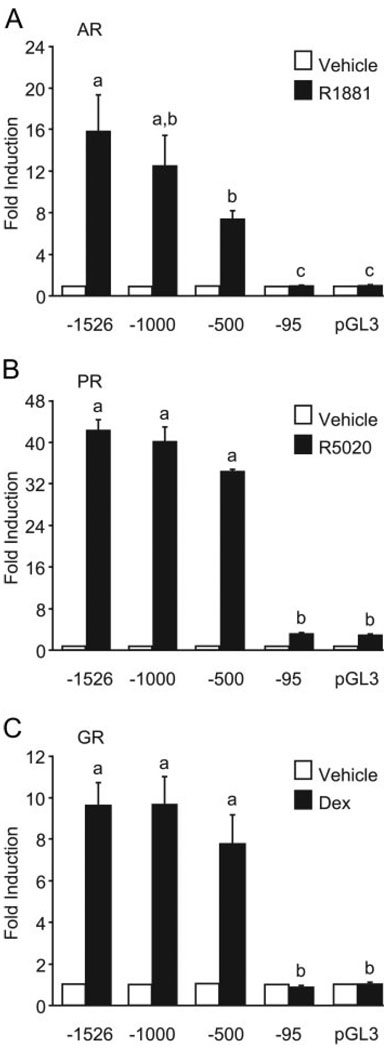
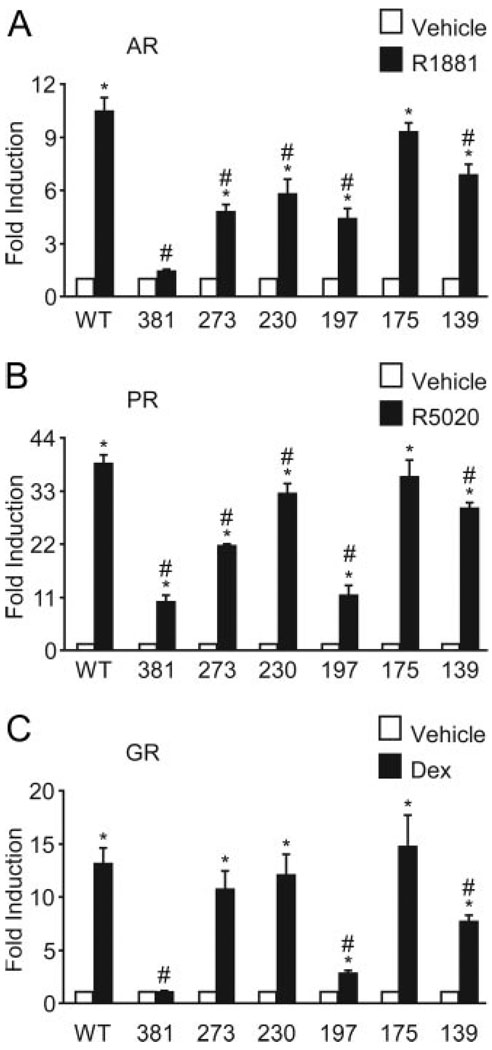
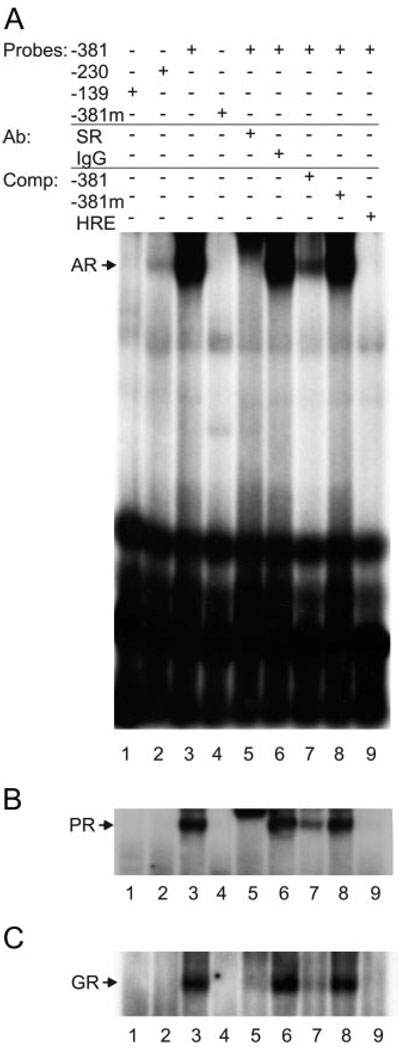
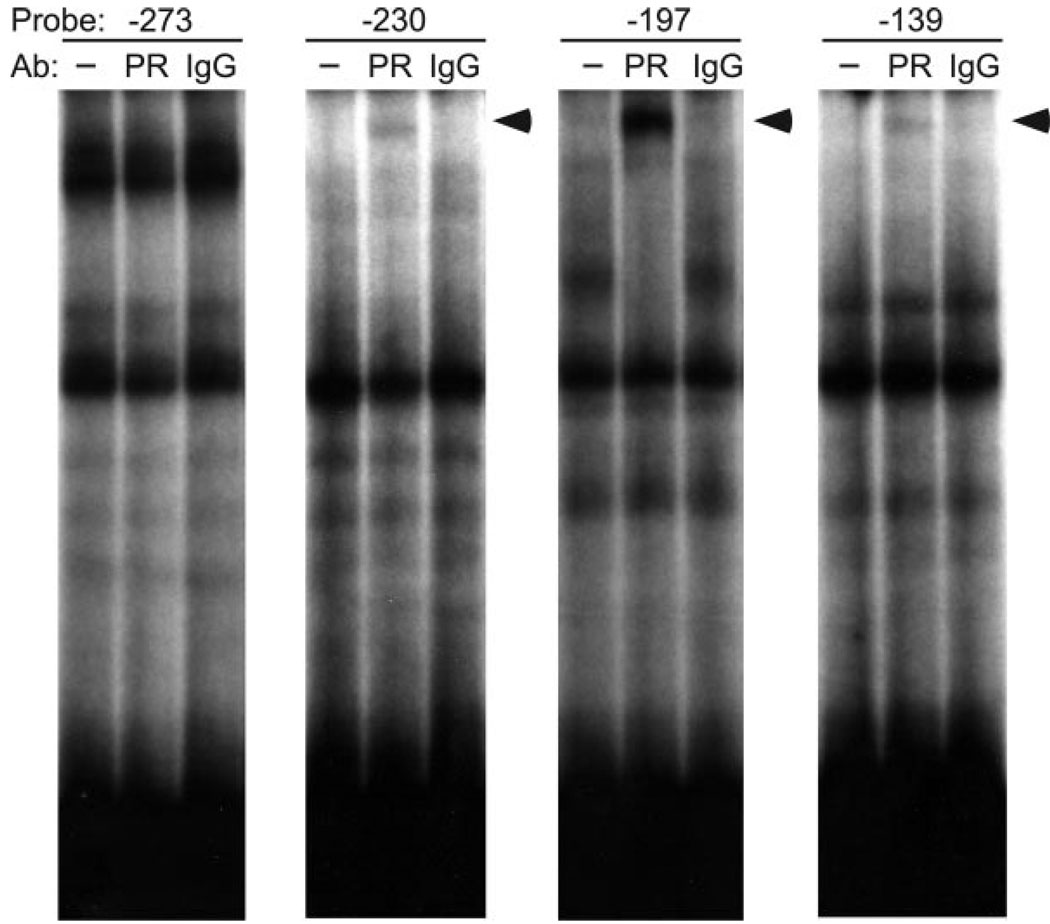
FSH is produced by the pituitary gonadotrope to regulate gametogenesis. Steroid hormones, including androgens, progestins, and glucocorticoids, have all been shown to stimulate expression of the FSHbeta subunit in primary pituitary cells and rodent models. Understanding the molecular mechanisms of steroid induction of FSHbeta has been difficult due to the heterogeneity of the anterior pituitary. Immortalized LbetaT2 cells are a model of a mature gonadotrope cell and express the endogenous steroid receptor for each of the three hormones. Transient transfection of each receptor, along with ligand treatment, stimulates the mouse FSHbeta promoter, but induction is severely diminished using receptors that lack the ability to bind DNA, indicating that induction is likely through direct DNA binding. All three steroid hormones act within the first 500 bp of the FSHbeta promoter where six putative hormone response elements exist. The -381 site is critical for FSHbeta induction by all three steroid hormones, whereas the -197 and -139 sites contribute to maximal induction. Interestingly, the -273 and -230 sites are also necessary for androgen and progestin induction of FSHbeta, but not for glucocorticoid induction. Additionally, we find that all three receptors bind the endogenous FSHbeta promoter, in vivo, and specifically bind the -381 site in vitro, suggesting that the binding of the receptors to this element is critical for the induction of FSHbeta by these 3-keto steroid hormones. Our data indicate that androgens, glucocorticoids, and progestins act via their receptors to directly activate FSHbeta gene expression in the pituitary gonadotrope.
Figures


Similar articles
-
Activin and glucocorticoids synergistically activate follicle-stimulating hormone beta-subunit gene expression in the immortalized LbetaT2 gonadotrope cell line.Endocrinology. 2007 Feb;148(2):762-73. doi: 10.1210/en.2006-0952. Epub 2006 Nov 2. Endocrinology. 2007. PMID: 17082263 Free PMC article.
-
Synergistic induction of follicle-stimulating hormone beta-subunit gene expression by gonadal steroid hormone receptors and Smad proteins.Endocrinology. 2008 Mar;149(3):1091-102. doi: 10.1210/en.2007-1498. Epub 2007 Dec 13. Endocrinology. 2008. PMID: 18079204 Free PMC article.
-
Cell-specific transcriptional regulation of follicle-stimulating hormone-beta by activin and gonadotropin-releasing hormone in the LbetaT2 pituitary gonadotrope cell model.Endocrinology. 2001 Jun;142(6):2284-95. doi: 10.1210/endo.142.6.8185. Endocrinology. 2001. PMID: 11356674
-
Regulation of gonadotropin subunit genes in tilapia.Comp Biochem Physiol B Biochem Mol Biol. 2001 Jun;129(2-3):489-502. doi: 10.1016/s1096-4959(01)00345-1. Comp Biochem Physiol B Biochem Mol Biol. 2001. PMID: 11399484 Review.
-
Overlapping but distinct profiles of gene expression elicited by glucocorticoids and progestins.Recent Prog Horm Res. 2003;58:199-226. doi: 10.1210/rp.58.1.199. Recent Prog Horm Res. 2003. PMID: 12795420 Review.
Cited by
-
Interaction between gonadotropin-releasing hormone and bone morphogenetic protein-6 and -7 signaling in LβT2 gonadotrope cells.Mol Cell Endocrinol. 2012 Jan 2;348(1):147-54. doi: 10.1016/j.mce.2011.08.001. Epub 2011 Aug 9. Mol Cell Endocrinol. 2012. PMID: 21846488 Free PMC article.
-
Runt-related transcription factors impair activin induction of the follicle-stimulating hormone {beta}-subunit gene.Endocrinology. 2010 Jun;151(6):2669-80. doi: 10.1210/en.2009-0949. Epub 2010 Mar 31. Endocrinology. 2010. PMID: 20357224 Free PMC article.
-
β-catenin regulates GnRH-induced FSHβ gene expression.Mol Endocrinol. 2013 Feb;27(2):224-37. doi: 10.1210/me.2012-1310. Epub 2012 Dec 4. Mol Endocrinol. 2013. PMID: 23211523 Free PMC article.
-
Glucocorticoids induce human glycoprotein hormone alpha-subunit gene expression in the gonadotrope.Endocrinology. 2008 Jul;149(7):3643-55. doi: 10.1210/en.2007-1100. Epub 2008 Apr 10. Endocrinology. 2008. PMID: 18403486 Free PMC article.
-
Progesterone Inhibits basal and gonadotropin-releasing hormone induction of luteinizing hormone beta-subunit gene expression.Endocrinology. 2009 May;150(5):2395-403. doi: 10.1210/en.2008-1027. Epub 2008 Dec 23. Endocrinology. 2009. PMID: 19106225 Free PMC article.
References
-
- Kumar TR, Wang Y, Lu N, Matzuk MM. Follicle stimulating hormone is required for ovarian follicle maturation but not male fertility. Nat Genet. 1997;15:201–204. - PubMed
-
- Burns KH, Matzuk MM. Minireview: genetic models for the study of gonadotropin actions. Endocrinology. 2002;143:2823–2835. - PubMed
-
- Pierce JG, Parsons TF. Glycoprotein hormones: structure and function. Annu Rev Biochem. 1981;50:465–495. - PubMed
-
- Kaiser UB, Jakubowiak A, Steinberger A, Chin WW. Differential effects of gonadotropin-releasing hormone (GnRH) pulse frequency on gonadotropin subunit and GnRH receptor messenger ribonucleic acid levels in vitro. Endocrinology. 1997;138:1224–1231. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
