

Structure of the unusual seryl-tRNA synthetase reveals a distinct zinc-dependent mode of substrate recognition
- PMID: 16675947
- PMCID: PMC1478180
- DOI: 10.1038/sj.emboj.7601129
Structure of the unusual seryl-tRNA synthetase reveals a distinct zinc-dependent mode of substrate recognition
Abstract
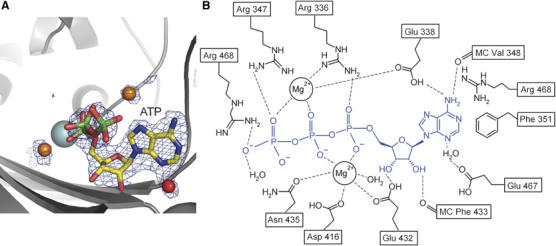
Methanogenic archaea possess unusual seryl-tRNA synthetase (SerRS), evolutionarily distinct from the SerRSs found in other archaea, eucaryotes and bacteria. The two types of SerRSs show only minimal sequence similarity, primarily within class II conserved motifs 1, 2 and 3. Here, we report a 2.5 A resolution crystal structure of the atypical methanogenic Methanosarcina barkeri SerRS and its complexes with ATP, serine and the nonhydrolysable seryl-adenylate analogue 5'-O-(N-serylsulfamoyl)adenosine. The structures reveal two idiosyncratic features of methanogenic SerRSs: a novel N-terminal tRNA-binding domain and an active site zinc ion. The tetra-coordinated Zn2+ ion is bound to three conserved protein ligands (Cys306, Glu355 and Cys461) and binds the amino group of the serine substrate. The absolute requirement of the metal ion for enzymatic activity was confirmed by mutational analysis of the direct zinc ion ligands. This zinc-dependent serine recognition mechanism differs fundamentally from the one employed by the bacterial-type SerRSs. Consequently, SerRS represents the only known aminoacyl-tRNA synthetase system that evolved two distinct mechanisms for the recognition of the same amino-acid substrate.
Figures





References
-
- Ahel D, Slade D, Mocibob M, Söll D, Weygand-Durasevic I (2005) Selective inhibition of divergent seryl-tRNA synthetases by serine analogues. FEBS Lett 579: 4344–4348 - PubMed
-
- Arnez JG, Dock-Bregeon AC, Moras D (1999) Glycyl-tRNA synthetase uses a negatively charged pit for specific recognition and activation of glycine. J Mol Biol 286: 1449–1459 - PubMed
-
- Baldwin AN, Berg P (1966) Transfer ribonucleic acid-induced hydrolysis of valyladenylate bound to isoleucyl ribonucleic acid synthetase. J Biol Chem 241: 839–845 - PubMed
-
- Baron B, Bock B (1995) The selenocysteine-inserting tRNA species: structure and function. In tRNA: Structure, Biosynthesis and Function, Söll D, RajBhandary U (eds), pp 529–544. Washington, DC: American Society for Microbiology
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
