

Displacement of D1, HP1 and topoisomerase II from satellite heterochromatin by a specific polyamide
- PMID: 16675949
- PMCID: PMC1478169
- DOI: 10.1038/sj.emboj.7601125
Displacement of D1, HP1 and topoisomerase II from satellite heterochromatin by a specific polyamide
Abstract
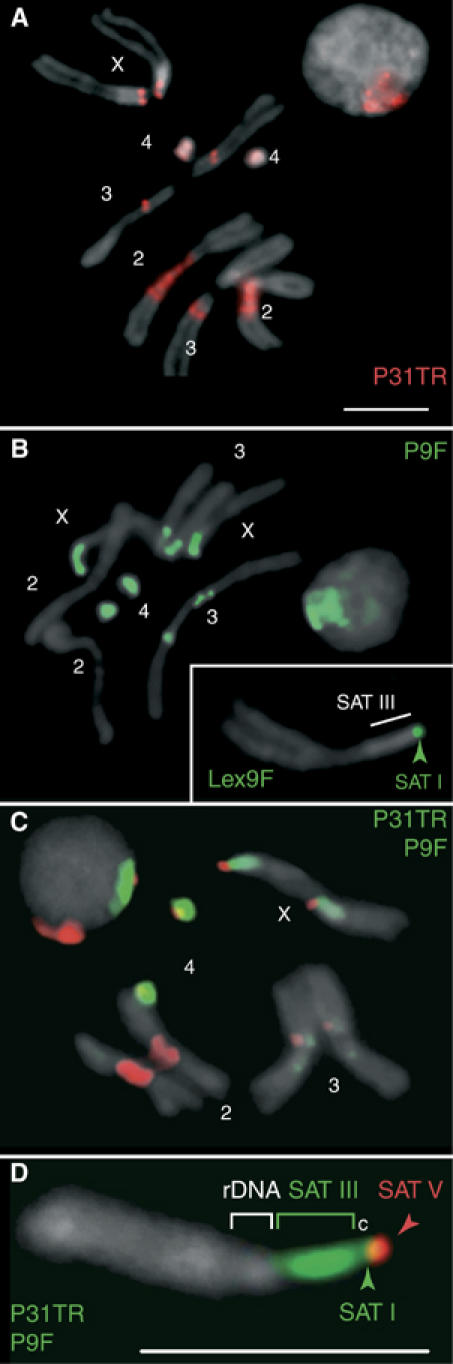
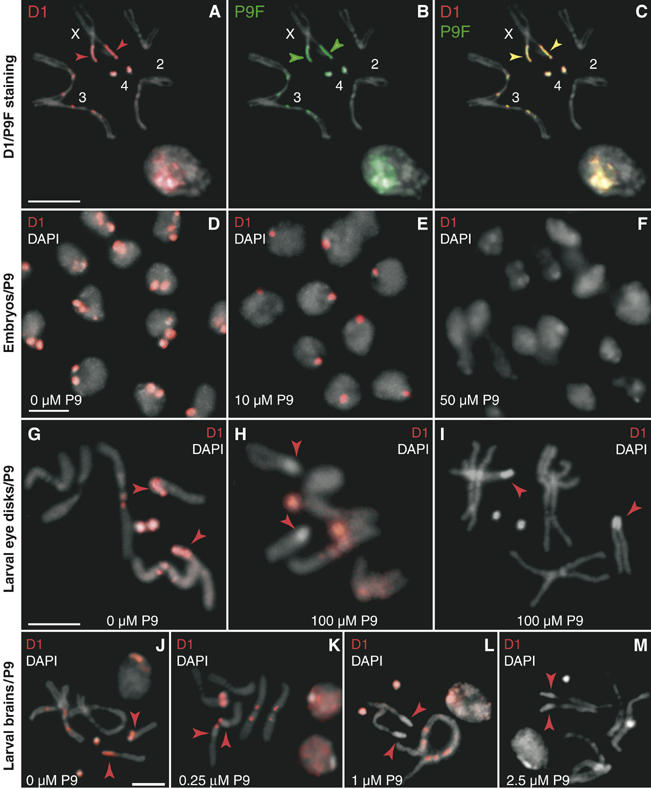
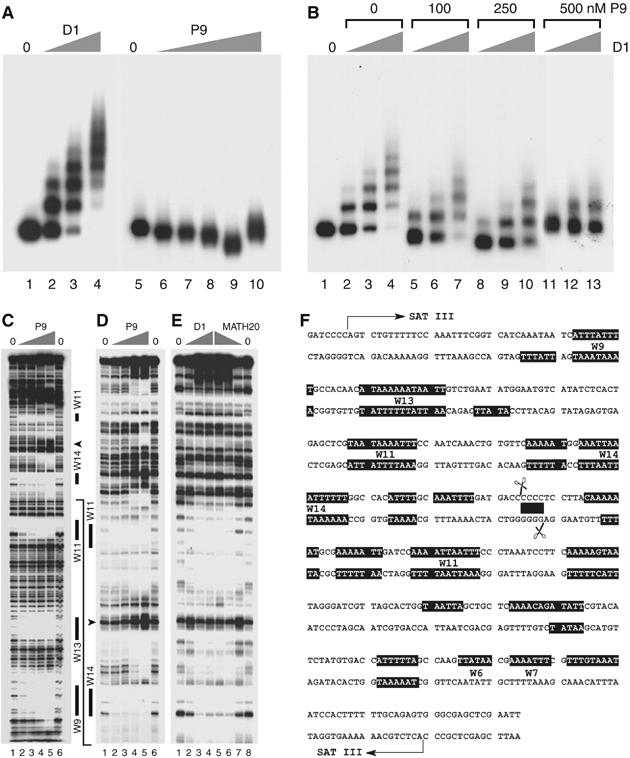
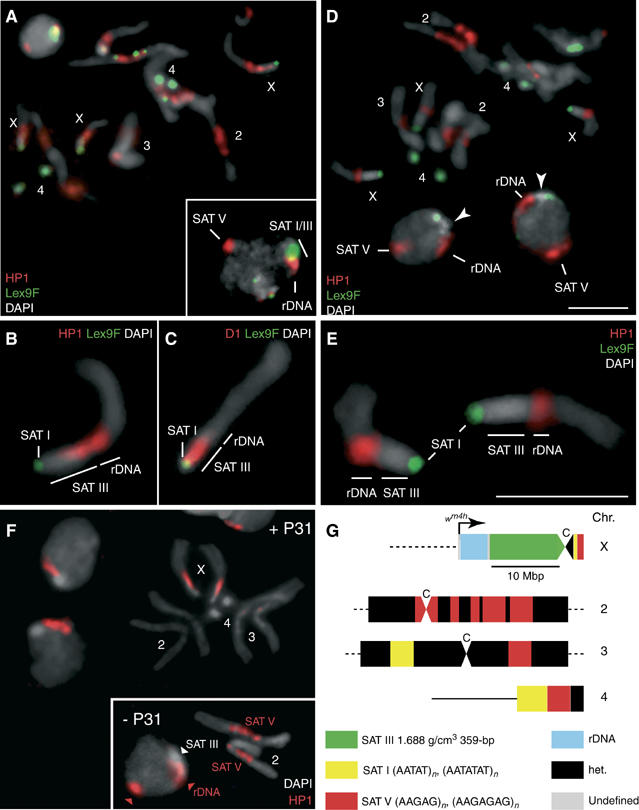
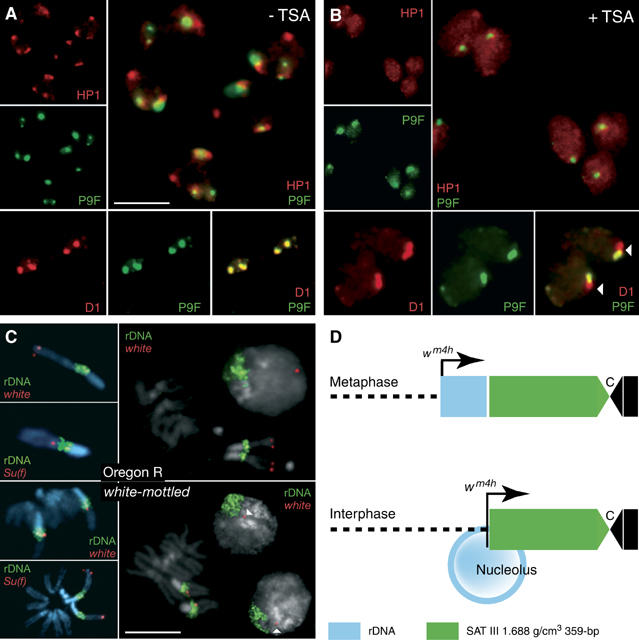
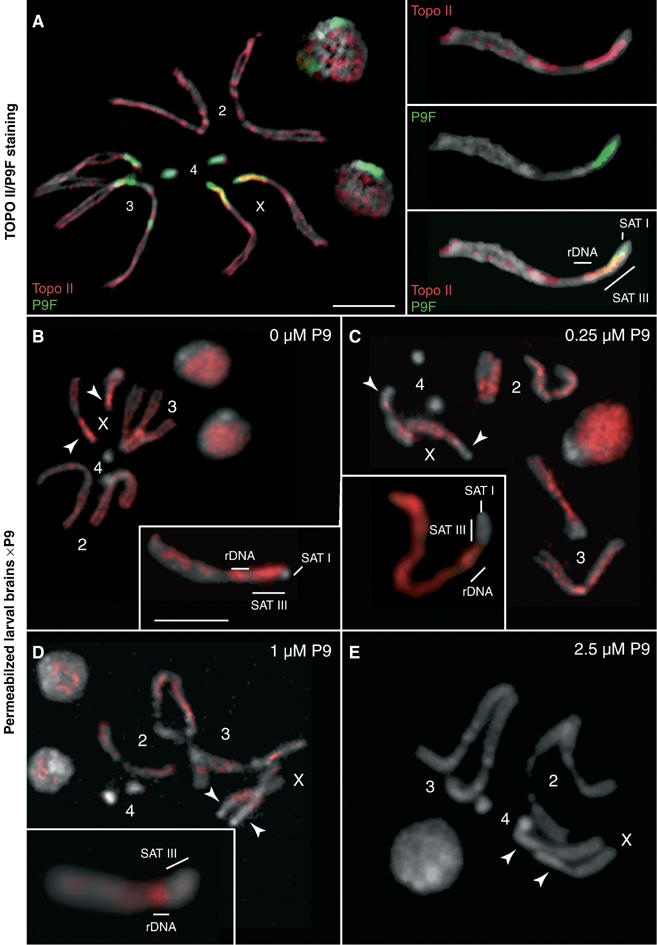
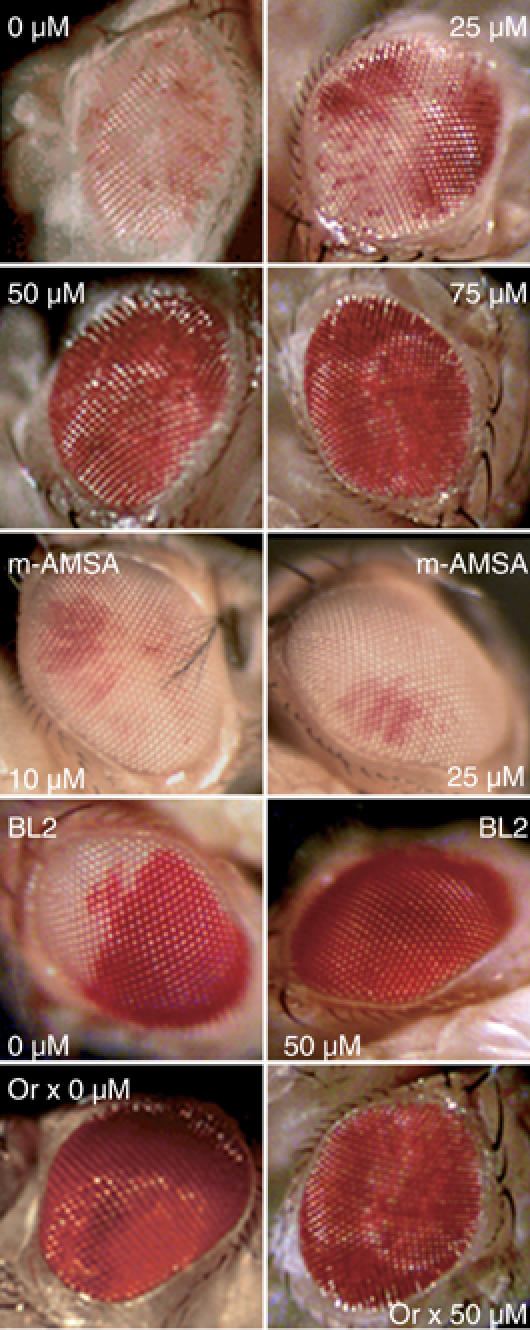
The functions of DNA satellites of centric heterochromatin are difficult to assess with classical molecular biology tools. Using a chemical approach, we demonstrate that synthetic polyamides that specifically target AT-rich satellite repeats of Drosophila melanogaster can be used to study the function of these sequences. The P9 polyamide, which binds the X-chromosome 1.688 g/cm3 satellite III (SAT III), displaces the D1 protein. This displacement in turn results in a selective loss of HP1 and topoisomerase II from SAT III, while these proteins remain bound to the adjacent rDNA repeats and to other regions not targeted by P9. Conversely, targeting of (AAGAG)n satellite V repeats by the P31 polyamide results in the displacement of HP1 from these sequences, indicating that HP1 interactions with chromatin are sensitive to DNA-binding ligands. P9 fed to larvae suppresses the position-effect variegation phenotype of white-mottled adult flies. We propose that this effect is due to displacement of the heterochromatin proteins D1, HP1 and topoisomerase II from SAT III, hence resulting in stochastic chromatin opening and desilencing of the nearby white gene.
Figures
References
-
- Adachi Y, Luke M, Laemmli UK (1991) Chromosome assembly in vitro: topoisomerase II is required for condensation. Cell 64: 137–148 - PubMed
-
- Ahmad K, Henikoff S (2002) The histone variant H3*3 marks active chromatin by replication-independent nucleosome assembly. Mol Cell 9: 1191–1200 - PubMed
-
- Aravin AA, Lagos-Quintana M, Yalcin A, Zavolan M, Marks D, Snyder B, Gaasterland T, Meyer J, Tuschl T (2003) The small RNA profile during Drosophila melanogaster development. Dev Cell 5: 337–350 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
