

PIASxbeta acts as an activator of Hoxb1 and is antagonized by Krox20 during hindbrain segmentation
- PMID: 16675951
- PMCID: PMC1478176
- DOI: 10.1038/sj.emboj.7601122
PIASxbeta acts as an activator of Hoxb1 and is antagonized by Krox20 during hindbrain segmentation
Abstract
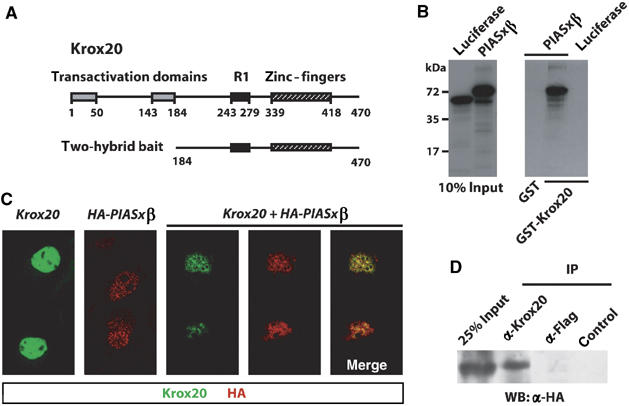
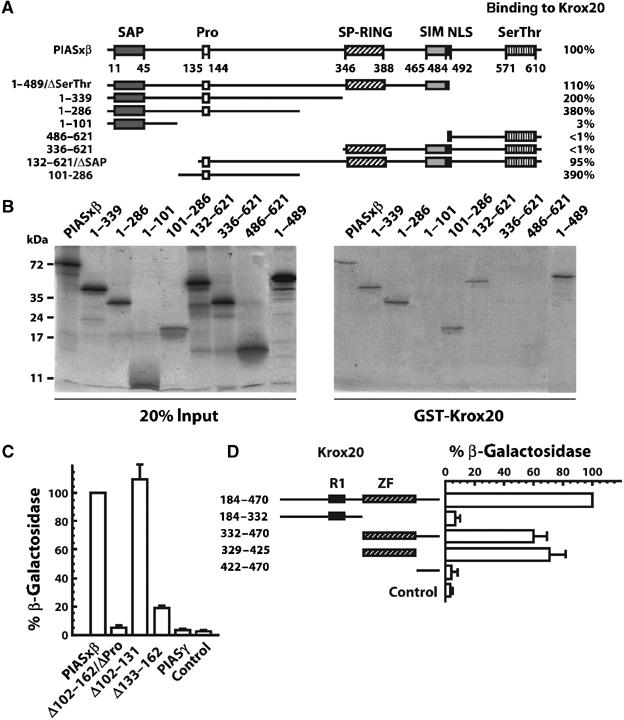
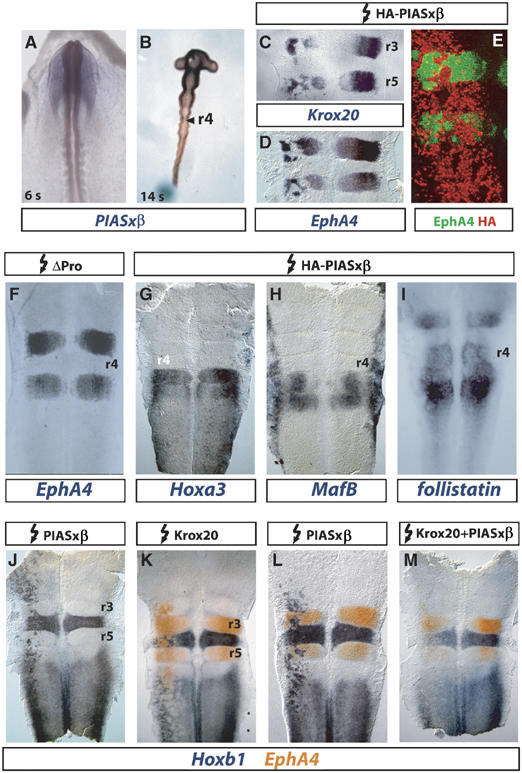
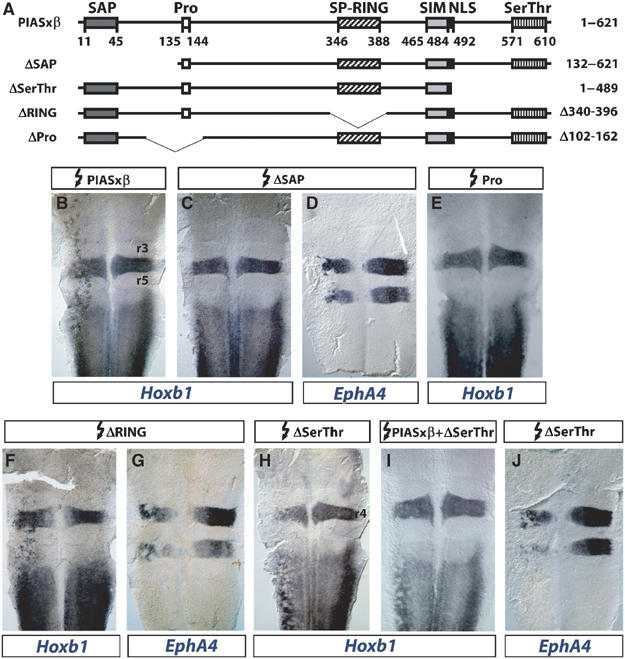
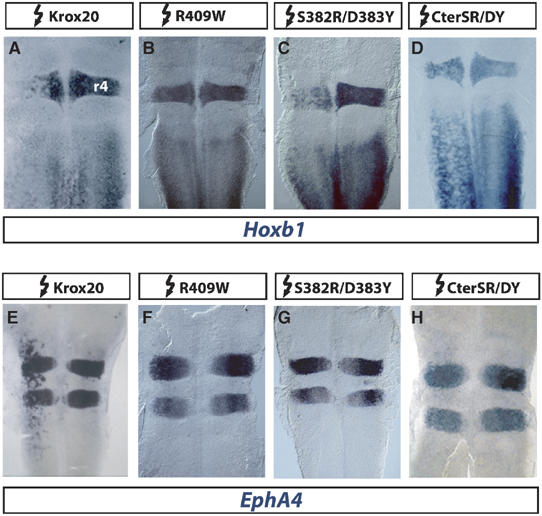
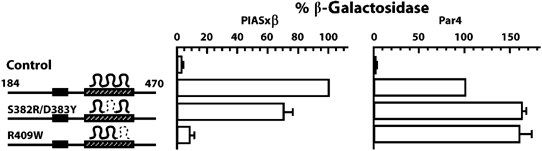
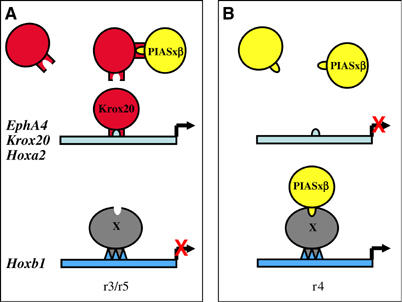
The zinc-finger transcription factor Krox20 constitutes a key regulator of hindbrain development, essential for the formation and specification of rhombomeres (r) 3 and 5. It is in particular responsible for the respective activation and repression of odd- and even-numbered rhombomere-specific genes, which include Hox genes. In this study, we have identified PIASxbeta as a novel direct interactor of Krox20. In addition, we found that PIASxbeta is able to activate the r4-specific gene Hoxb1. Binding of Krox20 prevents this activation, providing a molecular basis for the repression of Hoxb1 by Krox20. The same domain in the Krox20 protein, the zinc-fingers, is involved in DNA binding for transcriptional activation and in interaction with PIASxbeta for transcriptional repression, although the actual precise contacts are different. Our findings add an additional level in the complexity of Hox gene regulation and provide an example of how a single regulator can coordinate the activation and repression of a set of genes by very different mechanisms, acting as a molecular switch to specify cell identity and fate.
Figures
References
-
- Aravind L, Koonin EV (2000) SAP—a putative DNA-binding motif involved in chromosomal organization. Trends Biochem Sci 25: 112–114 - PubMed
-
- Barrow JR, Stadler HS, Capecchi MR (2000) Roles of Hoxa1 and Hoxa2 in patterning the early hindbrain of the mouse. Development 127: 933–944 - PubMed
-
- Birgbauer E, Sechrist J, Bronner-Fraser M, Fraser S (1995) Rhombomeric origin and rostrocaudal reassortment of neural crest cells revealed by intravital microscopy. Development 121: 935–945 - PubMed
-
- Cheng YC, Amoyel M, Qiu X, Jiang YJ, Xu Q, Wilkinson DG (2004) Notch activation regulates the segregation and differentiation of rhombomere boundary cells in the zebrafish hindbrain. Dev Cell 6: 539–550 - PubMed
-
- Choe SK, Sagerstrom CG (2004) Paralog group 1 hox genes regulate rhombomere 5/6 expression of vhnf1, a repressor of rostral hindbrain fates, in a meis-dependent manner. Dev Biol 271: 350–361 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
