

Overexpression of INFLORESCENCE DEFICIENT IN ABSCISSION activates cell separation in vestigial abscission zones in Arabidopsis
- PMID: 16679455
- PMCID: PMC1475485
- DOI: 10.1105/tpc.106.042036
Overexpression of INFLORESCENCE DEFICIENT IN ABSCISSION activates cell separation in vestigial abscission zones in Arabidopsis
Abstract
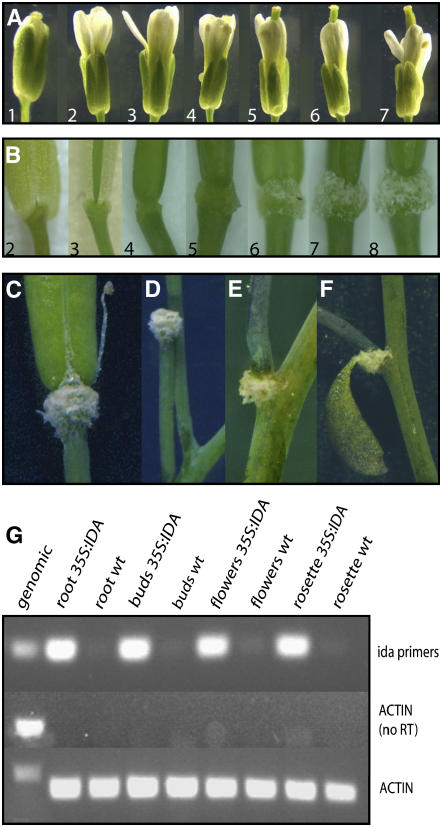
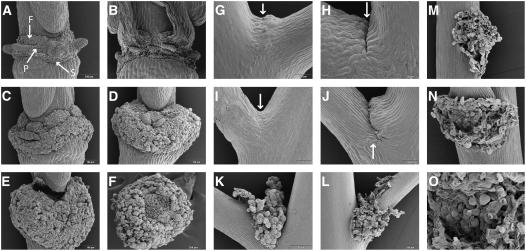
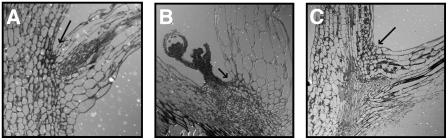
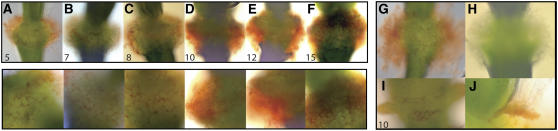
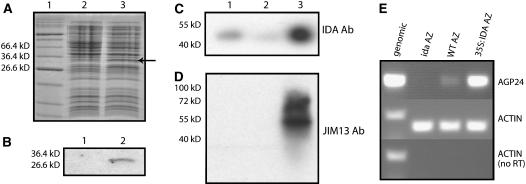
Plants may shed organs when they have been injured or served their purpose. The differential pattern of organ abscission in different species is most likely the result of evolutionary adaptation to a variety of life styles and environments. The final step of abscission-related cell separation in floral organs of wild-type Arabidopsis thaliana, which only abscises sepals, petals, and stamens, is controlled by INFLORESCENCE DEFICIENT IN ABSCISSION (IDA). Here, we demonstrate that Arabidopsis 35S:IDA lines constitutively overexpressing IDA exhibit earlier abscission of floral organs, showing that the abscission zones are responsive to IDA soon after the opening of the flowers. In addition, ectopic abscission was observed at the bases of the pedicel, branches of the inflorescence, and cauline leaves. The silique valves also dehisced prematurely. Scanning electron microscopy indicated a spread of middle lamella degradation from preformed abscission zone cells to neighboring cells. A transcript encoding an arabinogalactan protein (AGP) was upregulated in the 35S:IDA lines, and large amounts of AGP were secreted at the sites of abscission. AGP was shown to be a constituent of wild-type floral abscission zones during and soon after cell separation had been completed. We suggest that the restricted expression pattern of IDA precludes abscission of nonfloral organs in Arabidopsis.
Figures
References
-
- Aalen, R.B., Butenko, M.A., Stenvik, G.-E., Tandstad, N.M., and Patterson, S.E. (2006). Genetic control of floral abscission. In Floriculture, Ornamental and Plant Biotechnology: Advances and Topical Issues, Vol. 1, J.T. da Silva, ed (London: Global Science Books), pp. 101–108.
-
- Addicott, F.T. (1982). Abscission. (London: University of California Press).
-
- Alsop, D.W. (1974). Rapid single-solution polychrome staining of semithin epoxy sections using polyethylene glycol 200 (PEG 200) as a stain solvent. Stain Technol. 49 265–272. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
