

cAMP-Coupled riboflavin trafficking in placental trophoblasts: a dynamic and ordered process
- PMID: 16681382
- PMCID: PMC2525806
- DOI: 10.1021/bi060138f
cAMP-Coupled riboflavin trafficking in placental trophoblasts: a dynamic and ordered process
Abstract
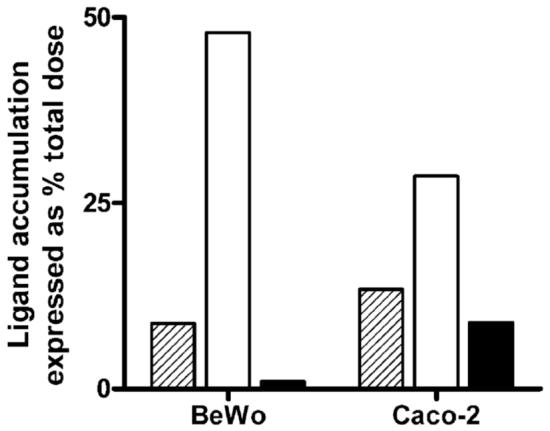
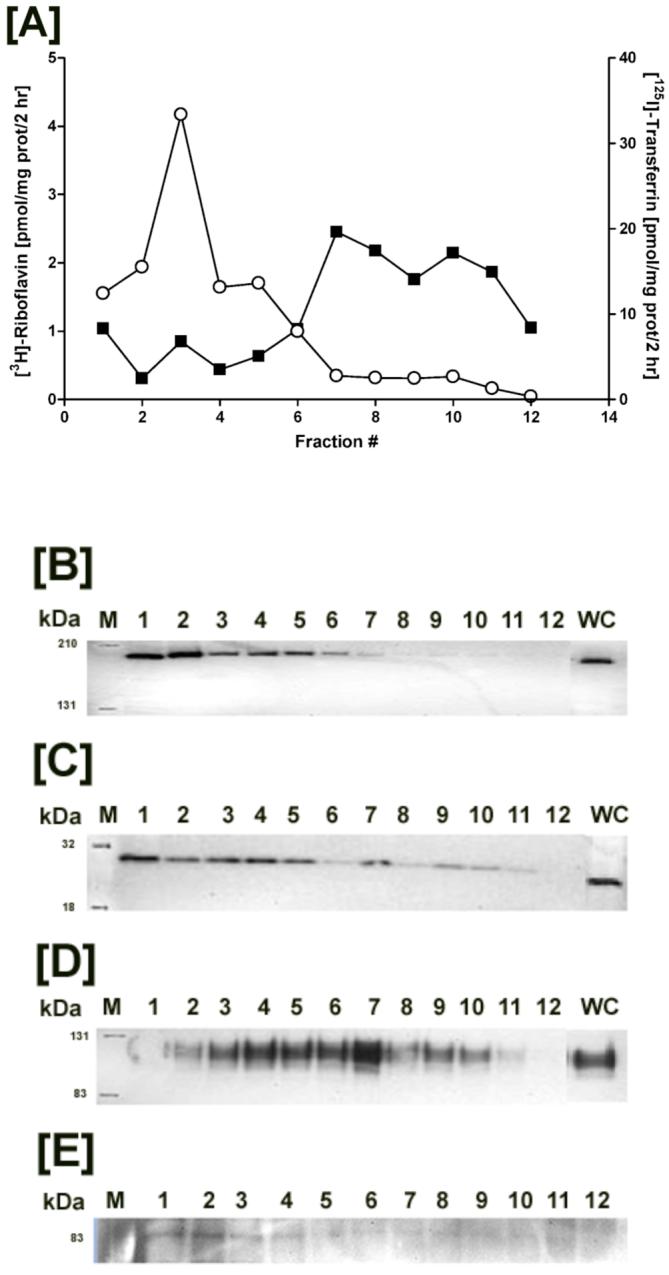
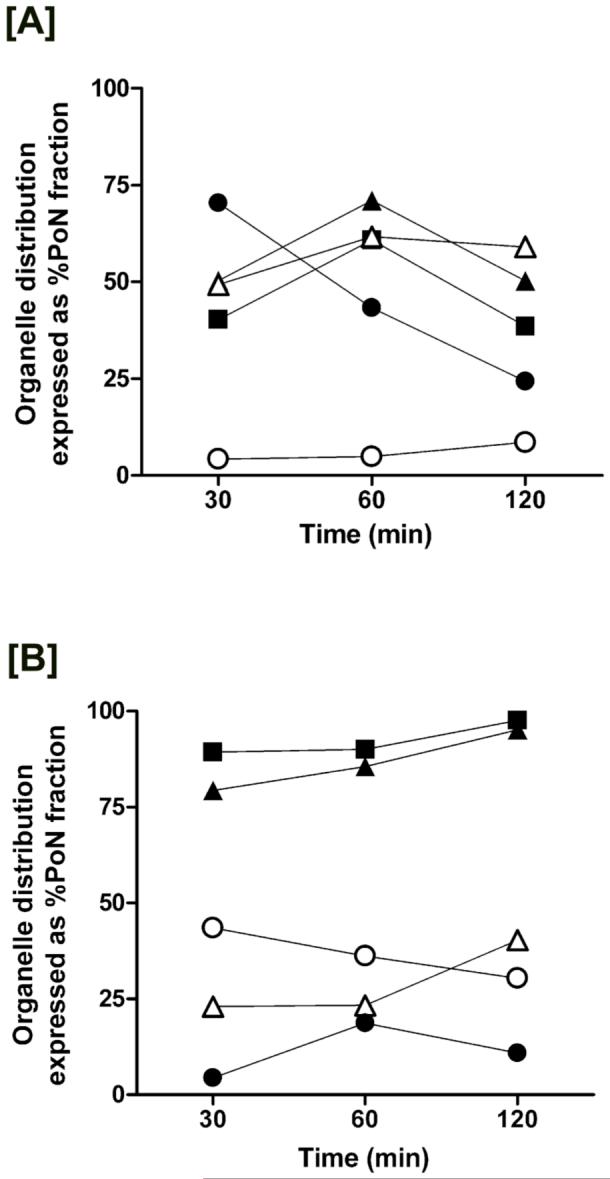
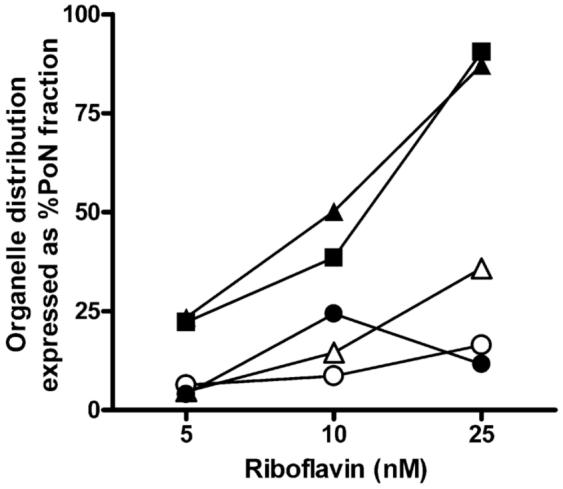
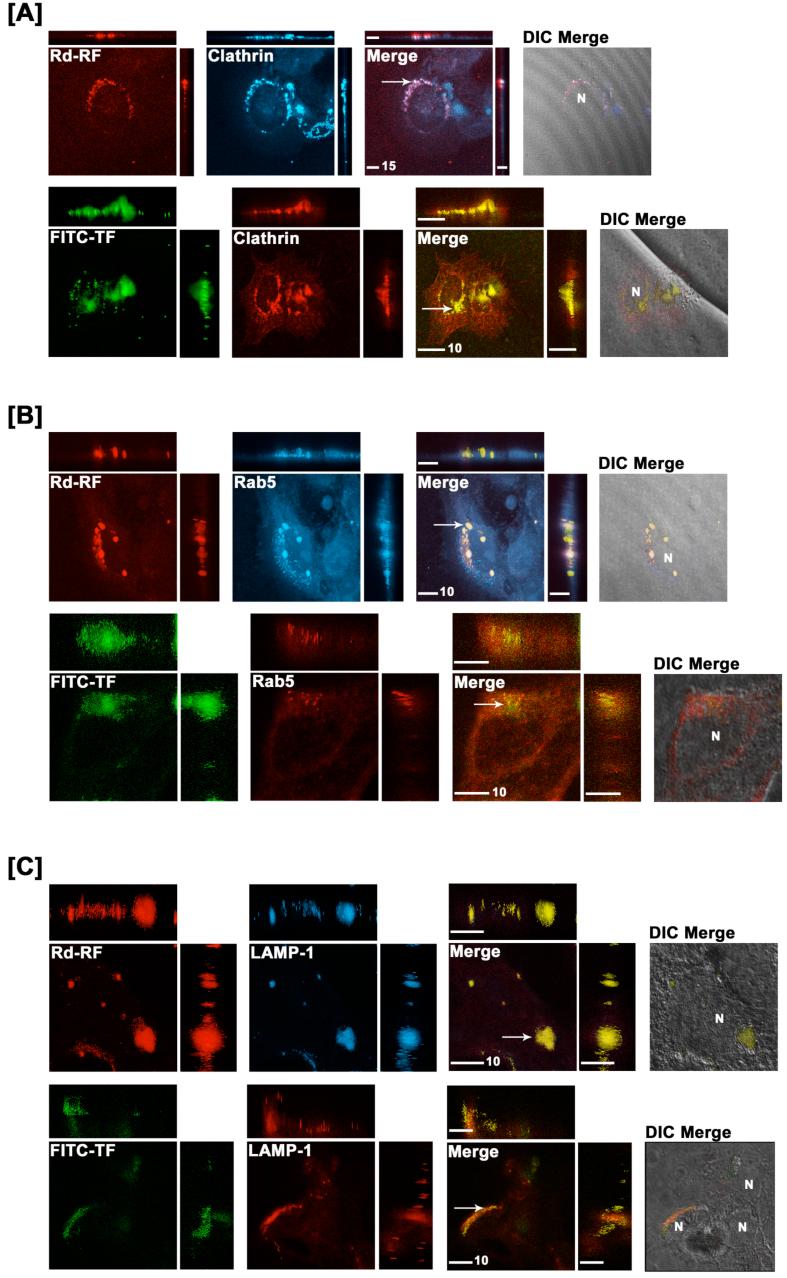
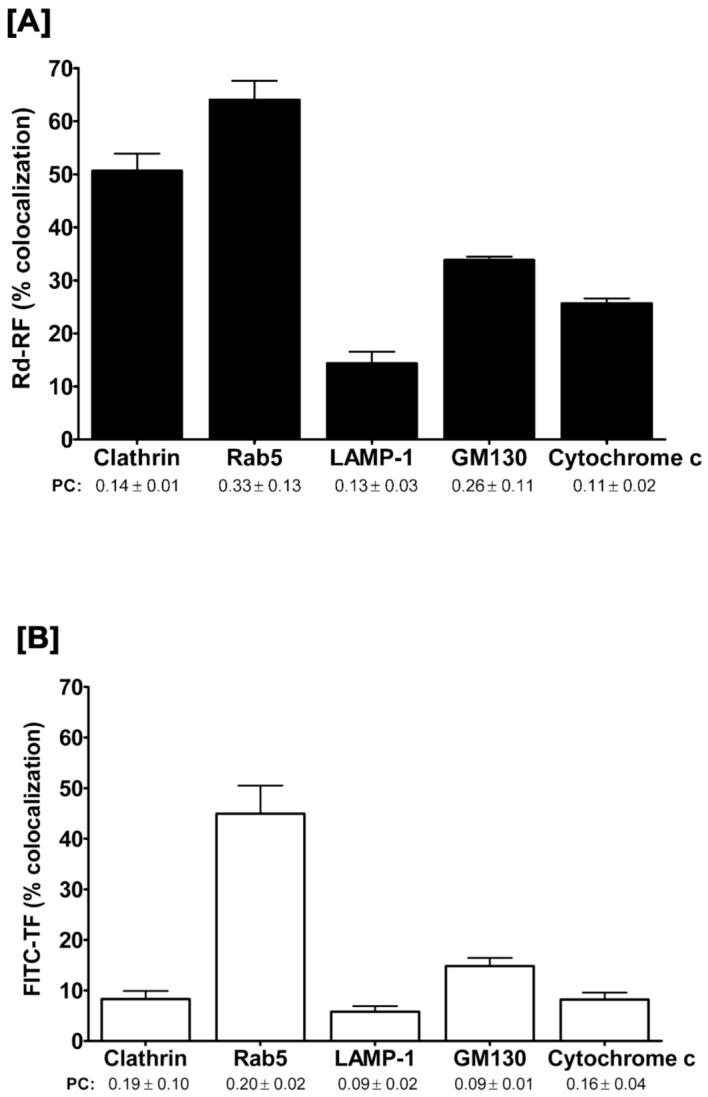
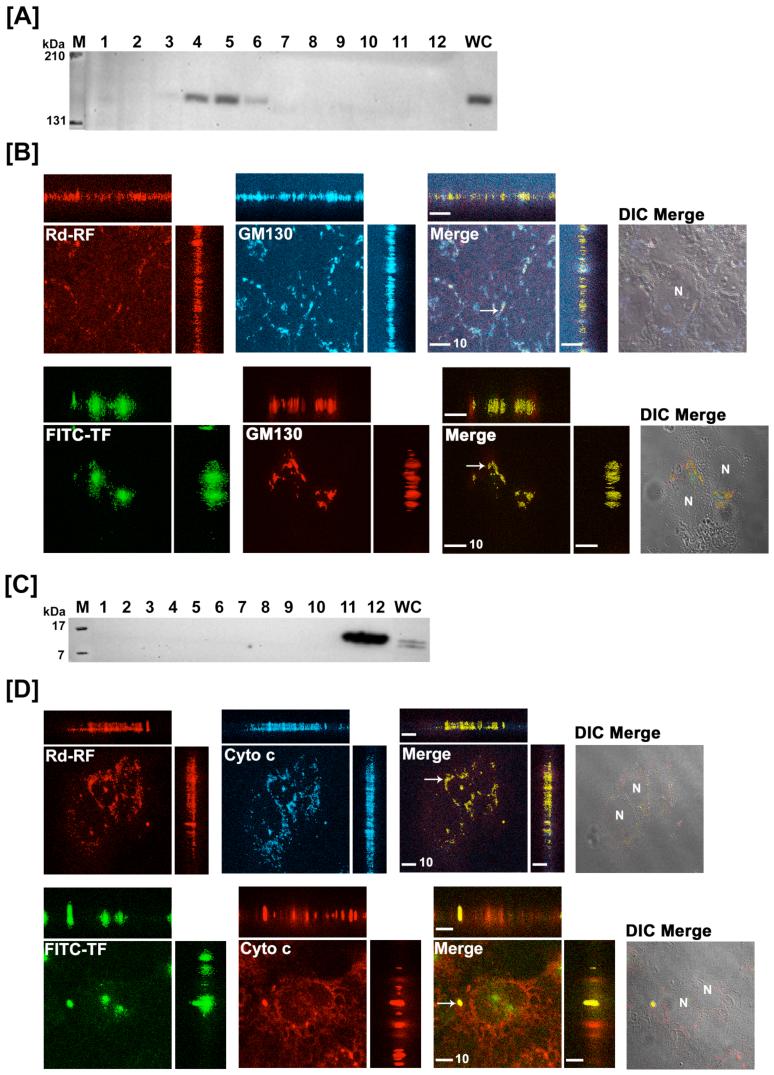
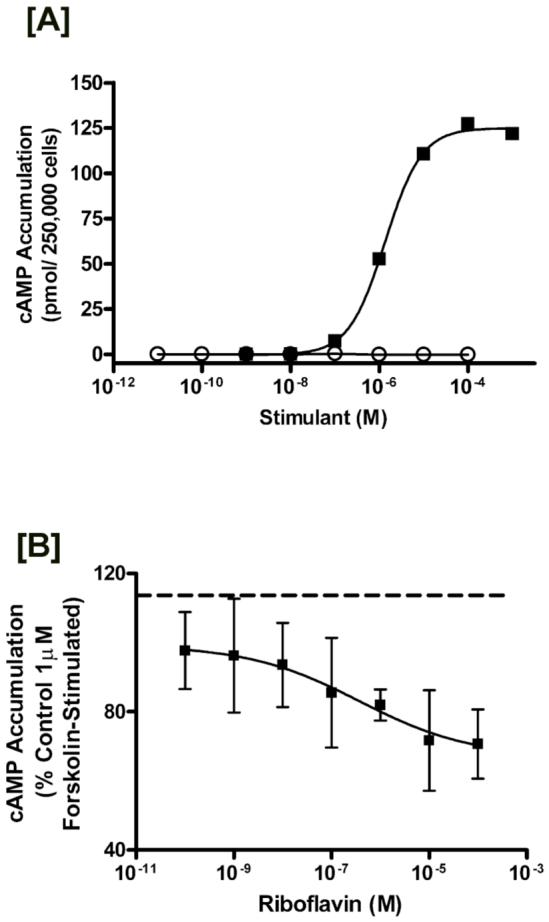
Riboflavin (RF, vitamin B(2)), an essential micronutrient central to cellular metabolism through formation of flavin mononucleotide (FMN) and flavin adenine dinucleotide (FAD) cofactors, is internalized, at least in part, via a proposed receptor-mediated endocytic (RME) process. The purpose of this study was to delineate the cellular RF distribution using human placental trophoblasts and evaluate the regulatory role of cAMP in this process. Subcellular fractionation and three-dimensional confocal microscopy analyses were carried out to define the RF accumulation profile. Biochemical assays evaluating the cAMP dependence of this pathway were also performed. This study records an intracellular RF distribution pattern that shows dynamic accumulation of the ligand predominantly in the endosomal and lysosomal compartments and to a lesser extent in the Golgi and mitochondria. In contrast, transferrin (TF) colocalizes rapidly within endosomes with minimal accumulation in the other organelles. The temporal and spatial distribution of RF and TF colocalized with unique markers of the endocytic machinery provides added morphological evidence in support of the RME process with ultimate translocation to the mitochondrial domain. Colocalized staining with the Golgi also suggests a possible recycling or exocytic mechanism for this ligand. Furthermore, this study demonstrates cAMP regulation of the putative ligand-bound RF receptor and its association into endocytic vesicles. Delineating the dynamics of the process governing cellular RF homeostasis presents an untapped resource that can be further exploited in improving our current understanding of nutritional biology and fetal growth and development, and perhaps in targeting the endogenous system for developing novel therapeutic approaches.
Figures
References
-
- Cooperman JM, Lopez R. Handbook of vitamins. Marcel Dekker; New York: 1991. Riboflavin; pp. 283–310.
-
- Powers HJ. Riboflavin (vitamin B-2) and health. Am. J. Clin. Nutr. 2003;77:1352–60. - PubMed
-
- Huang SN, Swaan PW. Involvement of a receptor-mediated component in cellular translocation of riboflavin. J. Pharmacol. Exp. Ther. 2000;294:117–25. - PubMed
-
- Huang SN, Swaan PW. Riboflavin uptake in human trophoblast-derived BeWo cell monolayers: cellular translocation and regulatory mechanisms. J. Pharmacol. Exp. Ther. 2001;298:264–71. - PubMed
-
- Foraker AB, Khantwal CM, Swaan PW. Current perspectives on the cellular uptake and trafficking of riboflavin. Adv. Drug Deliv. Rev. 2003;55:1467–83. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
