

Regulatory genes controlling fatty acid catabolism and peroxisomal functions in the filamentous fungus Aspergillus nidulans
- PMID: 16682457
- PMCID: PMC1459687
- DOI: 10.1128/EC.5.5.794-805.2006
Regulatory genes controlling fatty acid catabolism and peroxisomal functions in the filamentous fungus Aspergillus nidulans
Abstract
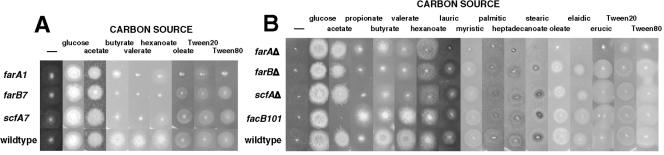
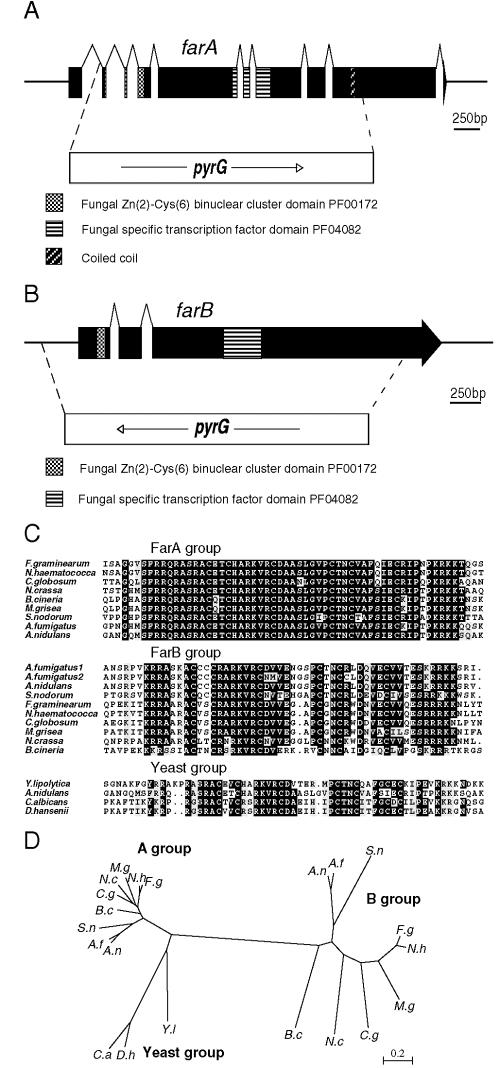
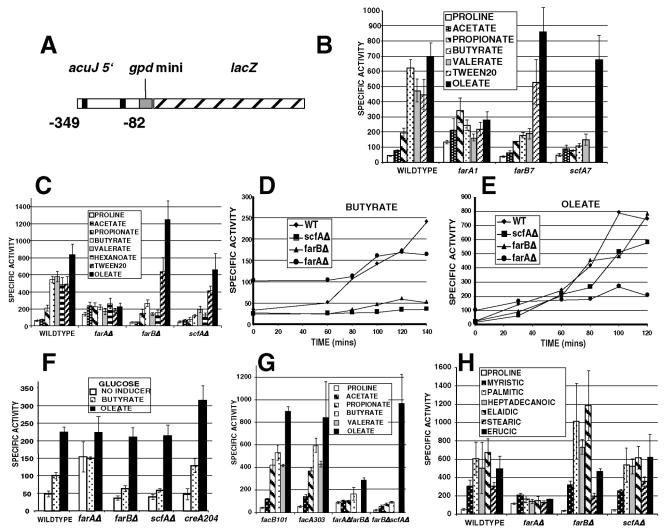
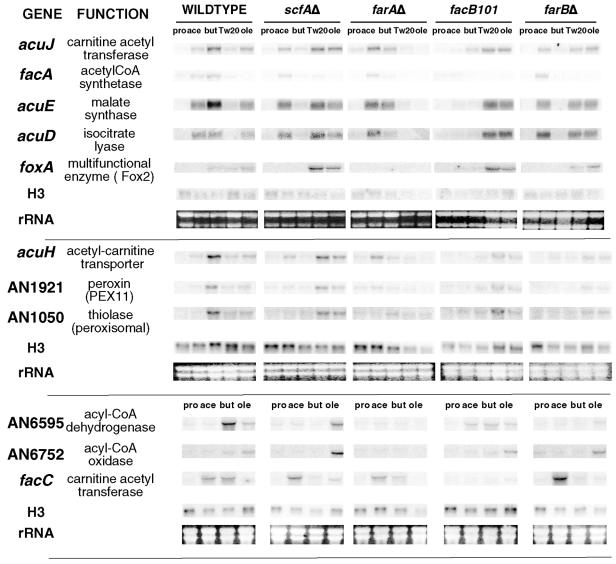
The catabolism of fatty acids is important in the lifestyle of many fungi, including plant and animal pathogens. This has been investigated in Aspergillus nidulans, which can grow on acetate and fatty acids as sources of carbon, resulting in the production of acetyl coenzyme A (CoA). Acetyl-CoA is metabolized via the glyoxalate bypass, located in peroxisomes, enabling gluconeogenesis. Acetate induction of enzymes specific for acetate utilization as well as glyoxalate bypass enzymes is via the Zn2-Cys6 binuclear cluster activator FacB. However, enzymes of the glyoxalate bypass as well as fatty acid beta-oxidation and peroxisomal proteins are also inducible by fatty acids. We have isolated mutants that cannot grow on fatty acids. Two of the corresponding genes, farA and farB, encode two highly conserved families of related Zn2-Cys6 binuclear proteins present in filamentous ascomycetes, including plant pathogens. A single ortholog is found in the yeasts Candida albicans, Debaryomyces hansenii, and Yarrowia lipolytica, but not in the Ashbya, Kluyveromyces, Saccharomyces lineage. Northern blot analysis has shown that deletion of the farA gene eliminates induction of a number of genes by both short- and long-chain fatty acids, while deletion of the farB gene eliminates short-chain induction. An identical core 6-bp in vitro binding site for each protein has been identified in genes encoding glyoxalate bypass, beta-oxidation, and peroxisomal functions. This sequence is overrepresented in the 5' region of genes predicted to be fatty acid induced in other filamentous ascomycetes, C. albicans, D. hansenii, and Y. lipolytica, but not in the corresponding genes in Saccharomyces cerevisiae.
Figures

Similar articles
-
An ortholog of farA of Aspergillus nidulans is implicated in the transcriptional activation of genes involved in fatty acid utilization in the yeast Yarrowia lipolytica.Biochem Biophys Res Commun. 2010 Nov 26;402(4):731-5. doi: 10.1016/j.bbrc.2010.10.096. Epub 2010 Oct 25. Biochem Biophys Res Commun. 2010. PMID: 20977884
-
Genetic analysis of the role of peroxisomes in the utilization of acetate and fatty acids in Aspergillus nidulans.Genetics. 2008 Mar;178(3):1355-69. doi: 10.1534/genetics.107.085795. Epub 2008 Feb 3. Genetics. 2008. PMID: 18245820 Free PMC article.
-
AoxA is a major peroxisomal long chain fatty acyl-CoA oxidase required for beta-oxidation in A. nidulans.Curr Genet. 2010 Apr;56(2):139-50. doi: 10.1007/s00294-009-0286-2. Epub 2009 Dec 31. Curr Genet. 2010. PMID: 20043225
-
Characterization of the Far Transcription Factor Family in Aspergillus flavus.G3 (Bethesda). 2016 Oct 13;6(10):3269-3281. doi: 10.1534/g3.116.032466. G3 (Bethesda). 2016. PMID: 27534569 Free PMC article.
-
Ethanol catabolism in Aspergillus nidulans: a model system for studying gene regulation.Prog Nucleic Acid Res Mol Biol. 2001;69:149-204. doi: 10.1016/s0079-6603(01)69047-0. Prog Nucleic Acid Res Mol Biol. 2001. PMID: 11550794 Review.
Cited by
-
pH effect on strain-specific transcriptomes of the take-all fungus.PLoS One. 2020 Jul 30;15(7):e0236429. doi: 10.1371/journal.pone.0236429. eCollection 2020. PLoS One. 2020. PMID: 32730288 Free PMC article.
-
Roles of Peroxisomes in the Rice Blast Fungus.Biomed Res Int. 2016;2016:9343417. doi: 10.1155/2016/9343417. Epub 2016 Aug 16. Biomed Res Int. 2016. PMID: 27610388 Free PMC article. Review.
-
The transcription factor homolog CTF1 regulates {beta}-oxidation in Candida albicans.Eukaryot Cell. 2009 Oct;8(10):1604-14. doi: 10.1128/EC.00206-09. Epub 2009 Aug 21. Eukaryot Cell. 2009. PMID: 19700635 Free PMC article.
-
Regulation of nutrient utilization in filamentous fungi.Appl Microbiol Biotechnol. 2023 Oct;107(19):5873-5898. doi: 10.1007/s00253-023-12680-4. Epub 2023 Aug 4. Appl Microbiol Biotechnol. 2023. PMID: 37540250 Free PMC article. Review.
-
Physiological role of Acyl coenzyme A synthetase homologs in lipid metabolism in Neurospora crassa.Eukaryot Cell. 2013 Sep;12(9):1244-57. doi: 10.1128/EC.00079-13. Epub 2013 Jul 19. Eukaryot Cell. 2013. PMID: 23873861 Free PMC article.
References
-
- Armitt, S., W. McCullough, and C. F. Roberts. 1976. Analysis of acetate non-utilizing (acu) mutants in Aspergillus nidulans. J. Gen. Microbiol. 92:263-282. - PubMed
-
- Ballance, D. J., and G. Turner. 1986. Gene cloning in Aspergillus nidulans: isolation of the isocitrate lyase gene (acuD). Mol. Gen. Genet. 202:271-275. - PubMed
-
- Baumgartner, U., B. Hamilton, M. Piskacek, H. Ruis, and H. Rottensteiner. 1999. Functional analysis of the Zn2Cys6 transcription factors Oaf1p and Pip2p. Different roles in fatty acid induction of beta-oxidation in Saccharomyces cerevisiae. J. Biol. Chem. 274:22208-22216. - PubMed
-
- Bowyer, P., J. R. De Lucas, and G. Turner. 1994. Regulation of the expression of the isocitrate lyase gene (acuD) of Aspergillus nidulans. Mol. Gen. Genet. 242:484-489. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
