

Pseudo-messenger RNA: phantoms of the transcriptome
- PMID: 16683022
- PMCID: PMC1449882
- DOI: 10.1371/journal.pgen.0020023
Pseudo-messenger RNA: phantoms of the transcriptome
Abstract
The mammalian transcriptome harbours shadowy entities that resist classification and analysis. In analogy with pseudogenes, we define pseudo-messenger RNA to be RNA molecules that resemble protein-coding mRNA, but cannot encode full-length proteins owing to disruptions of the reading frame. Using a rigorous computational pipeline, which rules out sequencing errors, we identify 10,679 pseudo-messenger RNAs (approximately half of which are transposon-associated) among the 102,801 FANTOM3 mouse cDNAs: just over 10% of the FANTOM3 transcriptome. These comprise not only transcribed pseudogenes, but also disrupted splice variants of otherwise protein-coding genes. Some may encode truncated proteins, only a minority of which appear subject to nonsense-mediated decay. The presence of an excess of transcripts whose only disruptions are opal stop codons suggests that there are more selenoproteins than currently estimated. We also describe compensatory frameshifts, where a segment of the gene has changed frame but remains translatable. In summary, we survey a large class of non-standard but potentially functional transcripts that are likely to encode genetic information and effect biological processes in novel ways. Many of these transcripts do not correspond cleanly to any identifiable object in the genome, implying fundamental limits to the goal of annotating all functional elements at the genome sequence level.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures
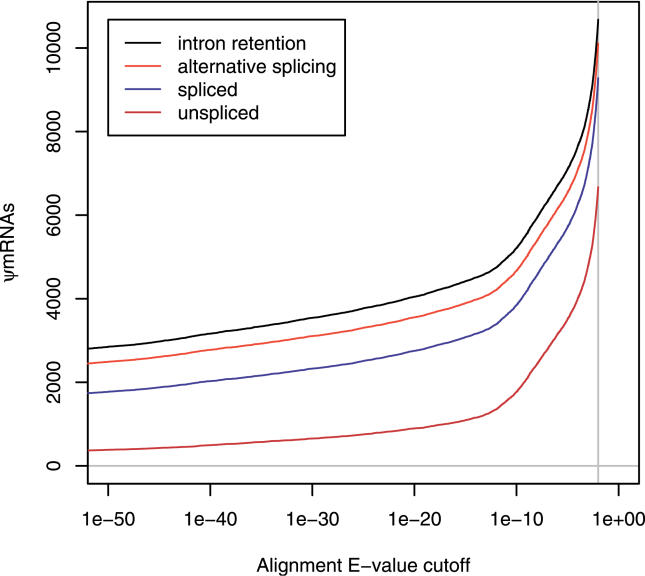



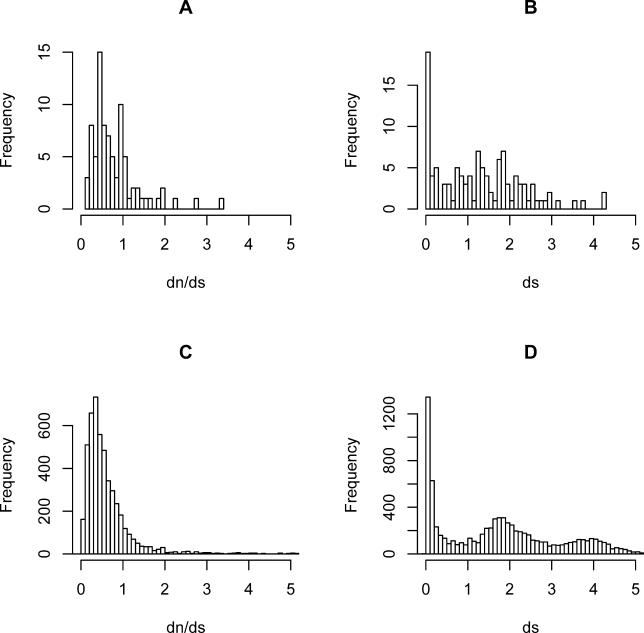


References
-
- Cheng J, Kapranov P, Drenkow J, Dike S, Brubaker S, et al. Transcriptional maps of 10 human chromosomes at 5-nucleotide resolution. Science. 2005;308:1149–1154. - PubMed
-
- Frith MC, Pheasant M, Mattick JS. Genomics: The amazing complexity of the human transcriptome. Eur J Hum Genet. 2005;13:894–897. - PubMed
-
- Carninci P, Kasukawa T, Katayama S, Gough J, Frith MC, et al. The transcriptional landscape of the mammalian genome. Science. 2005;309:1559–1563. - PubMed
-
- Mighell AJ, Smith NR, Robinson PA, Markham AF. Vertebrate pseudogenes. FEBS Lett. 2000;468:109–114. - PubMed
-
- Balakirev ES, Ayala FJ. Pseudogenes: Are they “junk” or functional DNA? Annu Rev Genet. 2003;37:123–151. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
