

Review
doi: 10.1021/cr040430y.
Early events in protein folding explored by rapid mixing methods
Affiliations
- PMID: 16683757
- PMCID: PMC2556641
- DOI: 10.1021/cr040430y
Item in Clipboard
Review
Early events in protein folding explored by rapid mixing methods
Chem Rev.
2006 May.
No abstract available
Figures
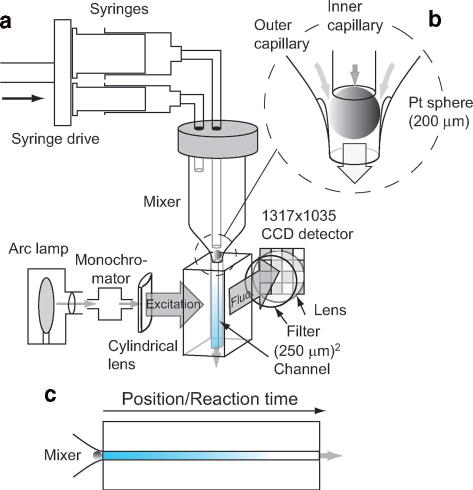
Continuous-flow capillary mixing apparatus in fluorescence mode. a Schematic of the solution delivery system, mixer, observation cell and optical arrangement. b Expanded view of the mixer. c Diagram illustrating continuous-flow measurement.
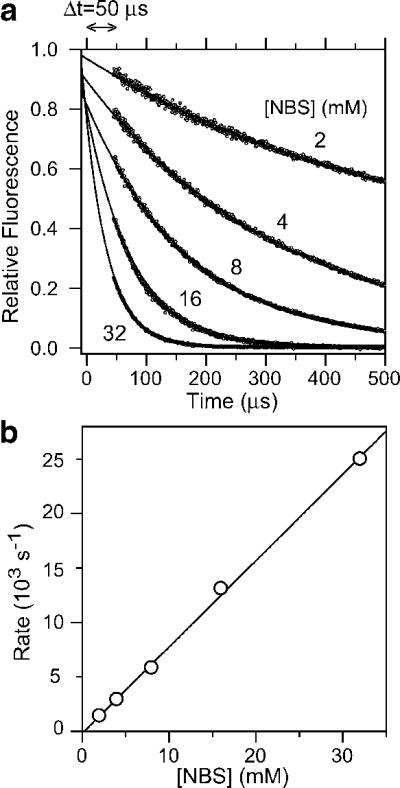
Continuous-flow measurements of the quenching of NATA fluorescence by NBS used to determine the experimental dead-time. a Plot of NATA fluorescence (>324 nm) vs. time at several NBS concentrations. b NATA-NBS reaction rates from exponential fitting of the data in panel a vs. NBS concentration. Linear regression (line) yields a second-order rate constant of 7.9 105 M−1 s−1.
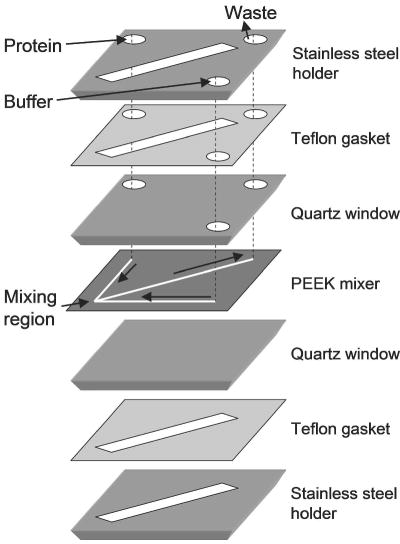
Schematic of a microfluidic mixer designed by Bilsel et al.. The 127-μm-thick mixer is sandwiched between quartz windows and is sealed in a stainless-steel holder using Teflon gaskets (1.6 mm thick). Solutions are delivered to and from the mixing region through holes in the upper layers (arrows). Reprinted with permission from ref . Copyright 2005 American Institute of Physics.
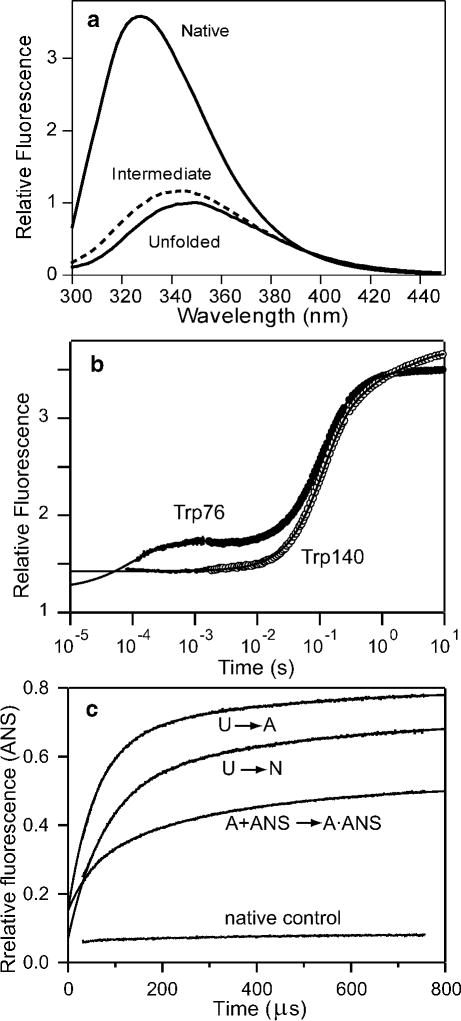
Folding mechanism of SNase probed by tryptophan fluorescence. a Fluorescence emission spectra of the Trp76 variant of SNase under native and denaturing conditions (solid) and a folding intermediate populated at equilibrium (dashed). The spectrum of the intermediate was determined by global analysis of the fluorescence spectra as a function of urea concentration (pH 5.2, 15 °C). b Time-course of folding (triggered by a pH jump from 2 to 5.2) for wild-type SNase (Trp140) and a single-tryptophan variant (Trp76) measured by continuous-flow (< 10−3 s) and stopped-flow (> 103 s) fluorescence. c ANS fluorescence changes during ANS binding/folding of Trp76 SNase measured by continuous-flow experiments at 15°C in the presence of 160 μM ANS. U → A: Salt concentration jump from 0 M to 1 M KCl at pH2.0; U → N: refolding induced by a pH-jump from 2.0 to 5.2; A+ANS → A•ANS: ANS binding kinetics in the presence of 1 M KCl at pH 2.0; native control: ANS binding kinetics under the native condition (pH 5.2). Adapted from ref (Figures 3 and 5).
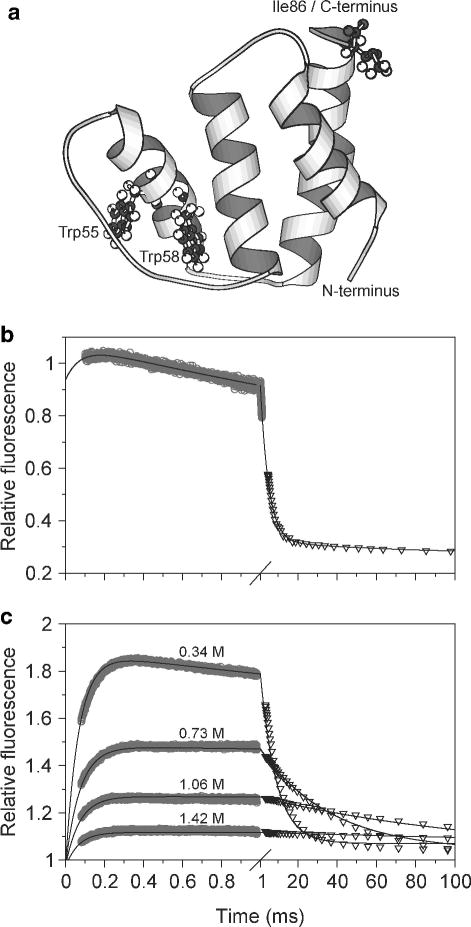
FRET-detection of an early folding intermediate in a helix-bundle protein, ACBP. a Ribbon diagram of ACBP, based on an NMR structure. The two tryptophan residues and the mutated C-terminal isoleucine are shown in ball and stick. The two lower panels show refolding kinetics of unmodified ACBP (b) and AEDANS-labeled ACBP,I86C (c) in pH 5.3 buffer containing 0.34 M GuHCl at 26 °C. In both panels data from continuous-flow (○) and stopped-flow (▽) experiments were matched and combined. Reprinted with permission from ref . Copyright 2002 National Academy of Sciences of the USA.
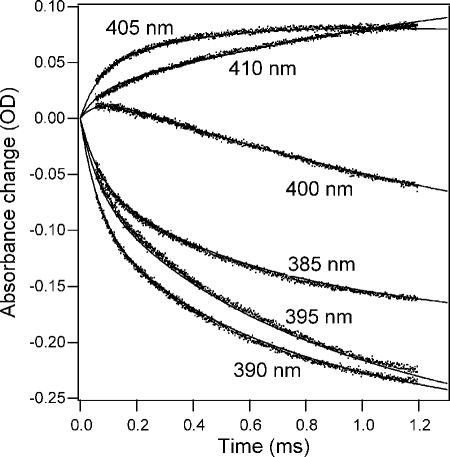
Initial stages of refolding of acid-denatured oxidized cyt c at pH 5 monitored by continuous-flow absorbance measurements at different wavelengths spanning the Soret heme absorbance band. The lines represent a global fit of a four-state folding mechanism to the family of kinetic traces. Reprinted with permission from ref . Copyright 2004 Elsevier Inc.
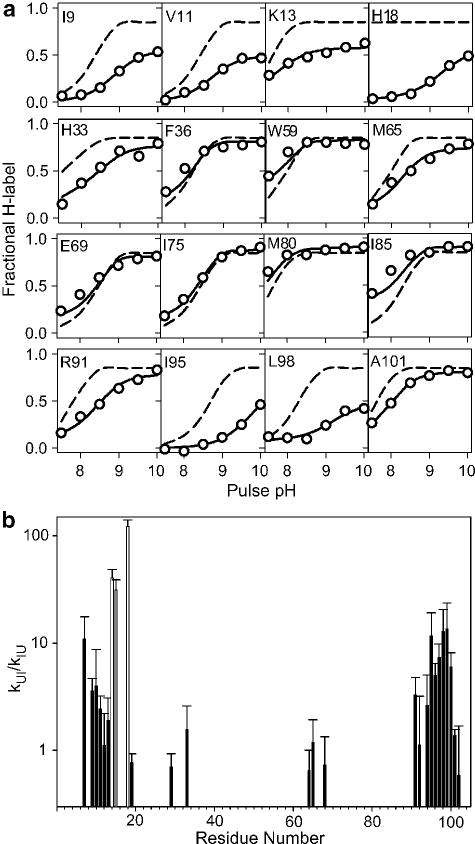
Characterization of an intermediate populated during folding of cytochrome c (pH 6, 10 °C). a Fractional degree of labeling vs. pulse pH for a representative set of NH groups measured by 2D NMR analysis of refolding cytochrome c samples that were exposed to a 50 ms labeling pulse of increasing pH at a folding time of 100 ms. Solid lines represent a fit of the data yielding the rates of formation and unfolding of the intermediate state (see text). Dashed lines indicate the labeling profiles expected in the absence of structure. b Equilibrium constant for formation of the intermediate, KUI = kUI/kIU, based on the fits of the labeling results in a. Values of KUI ≥ 1 indicative of persistent hydrogen bonded structure are mainly found for residues in the N- and C-terminal helices. Cys 14, Ala15 and His 18 (gray bars) are protected even in the unfolded state. Adapted from ref (Figures 3 and 4), with permission by the authors.
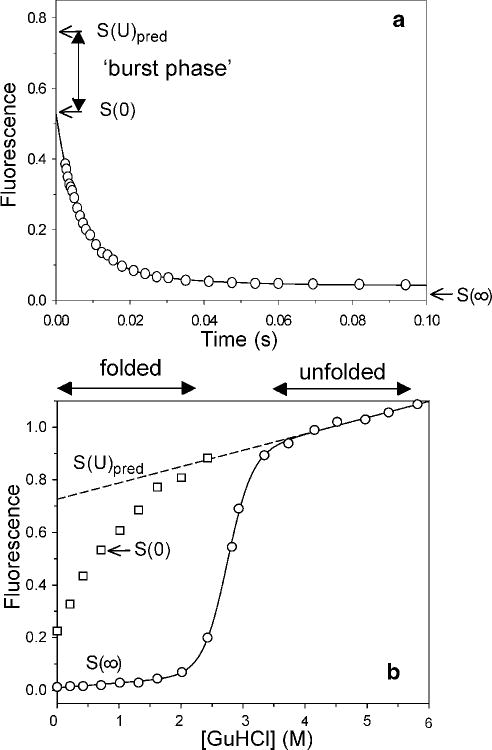
Stopped-flow fluorescence evidence for an unresolved rapid process (burst phase) during folding of cyt c (pH 5, 10 °C). a Tryptophan fluorescence changes during refolding of acid-unfolded cytochrome c (pH 2, ~15 mM HCl) at a final GuHCl concentration of 0.7 M. The initial signal S(0) at t=0 (determined on the basis of a separate dead-time measurement) falls short of the signal for the unfolded state under refolding conditions, Spred(U), obtained by linear extrapolation of the unfolded-state baseline (see dashed line in b). b Effect of the denaturant concentration on the initial (squares) and final (circles) fluorescence signal, S(0) and S(∞), measured in a series of stopped-flow refolding experiments at different final GuHCl concentration. Reprinted with permission from ref . Copyright 2004 Elsevier Inc.
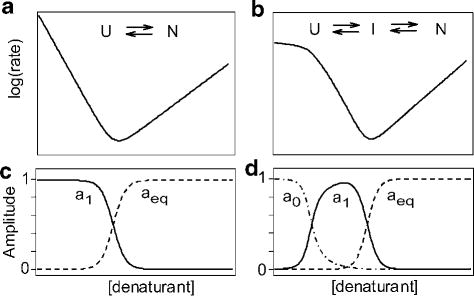
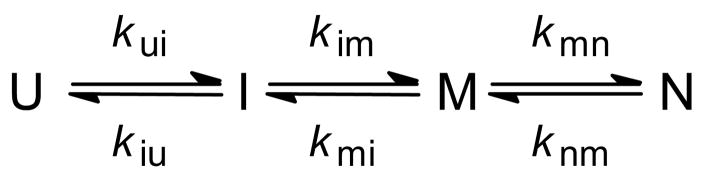
Schematic log(rate) vs. [denaturant] plots (chevrons) for a two-state (a) and a three-state (b) folding/unfolding mechanism. The lower panels show the predicted amplitude for the main folding phases (a1), the burst phase predicted for a three-state process (a0 in panel d) and the equilibrium unfolding transition (aeq).
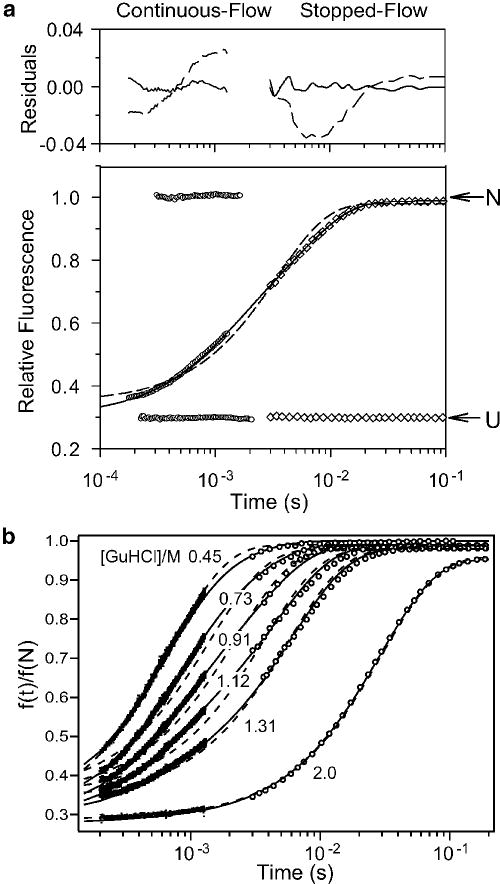
Folding kinetics of GB1 at pH 5.0, 20 °C, in the presence of 0.4 M sodium sulfate. Panel a shows a representative kinetic trace at 1.12 M GHCl monitored by continuous-flow (circles) and stopped-flow (diamonds) fluorescence along with controls for fully unfolded (U) and folded (N) solutions. Single- and double-exponential fits and residuals are shown with solid and dashed lines, respectively. Reprinted with permission from ref . Copyright 1999 Nature Publishing Group. Panel b shows a family of refolding traces at the final GuHCl concentrations indicated. Solid lines show the time course predicted on the basis of a three-state model (Scheme 3), using the following parameters: kUI° = 2300 s−1, mUI‡ = −0.6 kcal mol−1M−1, kIU° = 70 s−1, mIU‡ = 1.15 kcal mol−1M−1, kIN° = 600 s−1, mIN‡ = 0, kNI° = 0.14, mNI‡ = 0.3 kcal mol−1M−1 (see Figure 11a, ref 78). The relative signals for the N-, I- and U-states were sN = 0.29 ± 0.01, sI = sU = 0.98 ± 0.02. Dashed lines indicate a best “fit” of a two-state mechanism, using rate constants falling on a linear extrapolation of the chevron plot between 1.5 and 3 M GuHCl to lower denaturant concentrations (cf. ref 213).
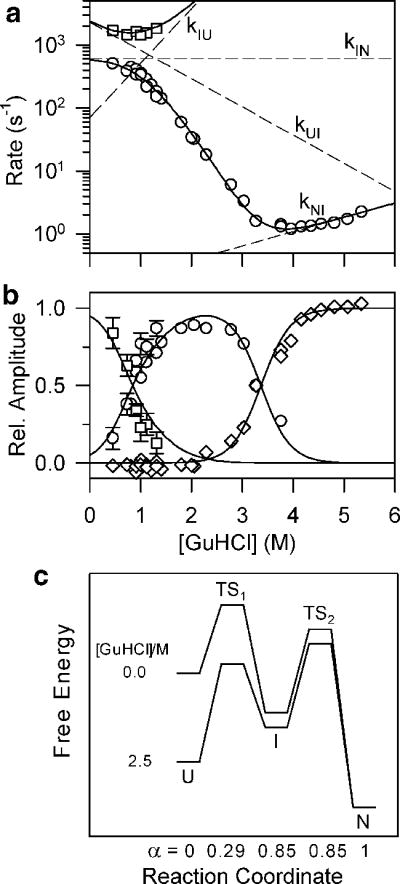
GuHCl-dependence of the rate constants (a) and kinetic amplitudes (b) of the fast (squares) and slow (circles) kinetic phases observed during folding of GB1. c Free energy diagrams for folding of GB1 under conditions where the intermediate, I is well populated (0 M) and unstable (2.5 M GuHCl). α represents the change in solvent-accessible surface area relative to the unfolded state U. Reprinted with permission from ref . Copyright 1999 Nature Publishing Group.
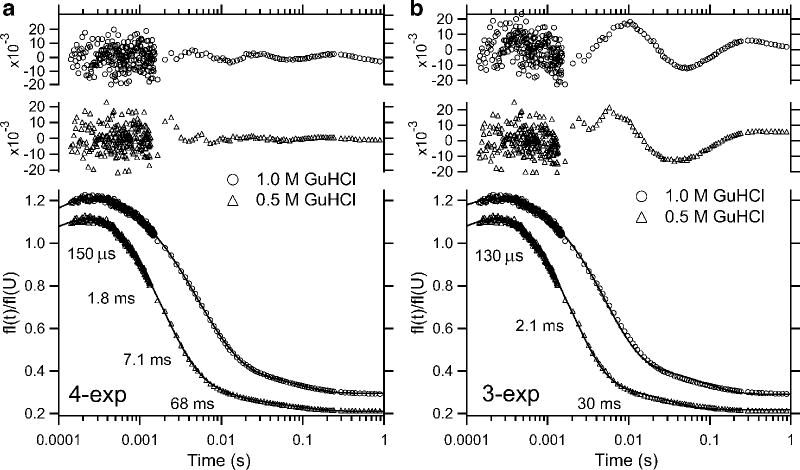
Comparison of quadruple (a) and triple (b) exponential fitting of the kinetics of refolding of F45W ubiquitin at final GuHCl concentrations of 0.5 and 1.0 M (pH 5, 25 °C). Fluorescence traces measured in continuous- and stopped-flow experiments were normalized with respect to the unfolded protein in 6 M GuHCl. The residuals (top two traces in each panel) indicate that four exponentials are required to obtain a satisfactory fit of the data over the time window shown. Reprinted with permission from ref . Copyright 2005 Wiley-VCH.
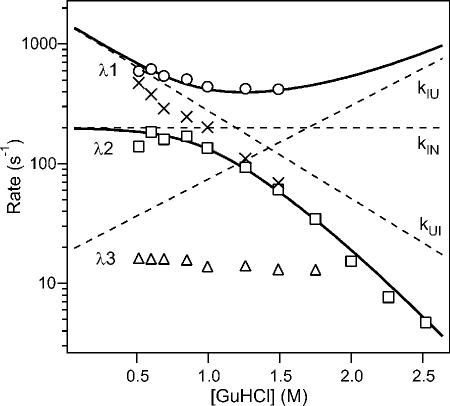
Expanded region of the rate profile (log rate vs. [GuHCl[) for the two main phases (circles and squares) and a minor slower phase (dtriangles) observed during folding of F45W ubiquitin (pH 5, 25 °C). The solid lines show the rates predicted by a three-state model, and the dashed lines indicate the elementary rate constants used. The X-symbols show the apparent rates obtained by triple-exponential fitting (Figure 10b). Reprinted with permission from ref . Copyright 2005 Wiley-VCH.
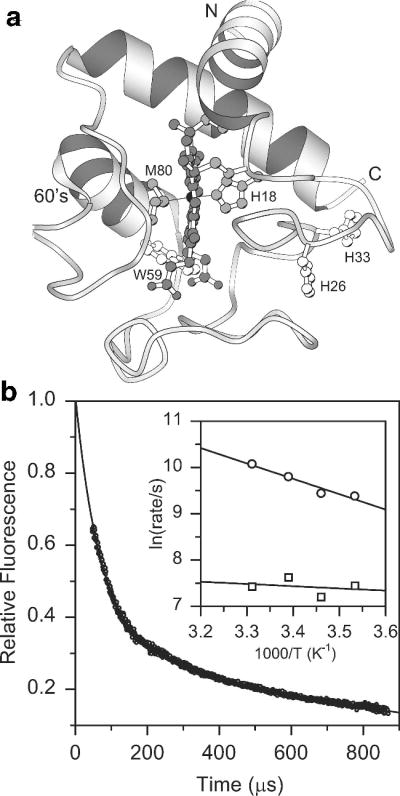
a Ribbon diagram of horse cytochrome c, based on the crystal structure. b Refolding kinetics of acid-unfolded cyt c at pH 4.5, 22 °C measured by continuous-flow mixing, indicating heme-induced quenching of Trp59 fluorescence associated with chain collapse. Inset: Arrhenius plot for the rates of the major (circles) and the minor (squares) submillisecond phases. Adapted from ref (Figure 2).
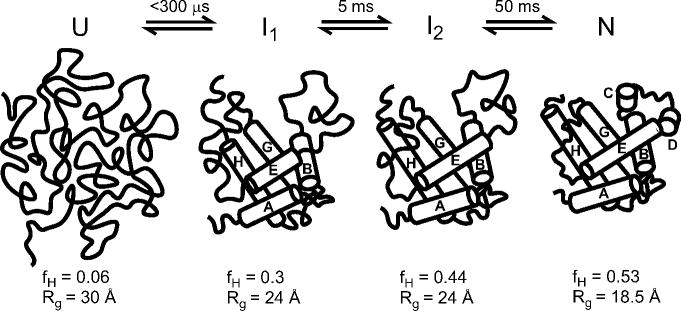
Sequential four-state mechanism involving two partially structured ensembles, I1 and I2, in addition to the acid-unfolded (U) and native (N) states. Cartoons of various states apomyoglobin consistent with the fraction of helical residues (fH) and radius of gyration (Rg) reported in ref . Cylinders indicate the approximate position of α-helices in the native state and possible arrangement of core helices in the intermediates consistent with amide protection data.,
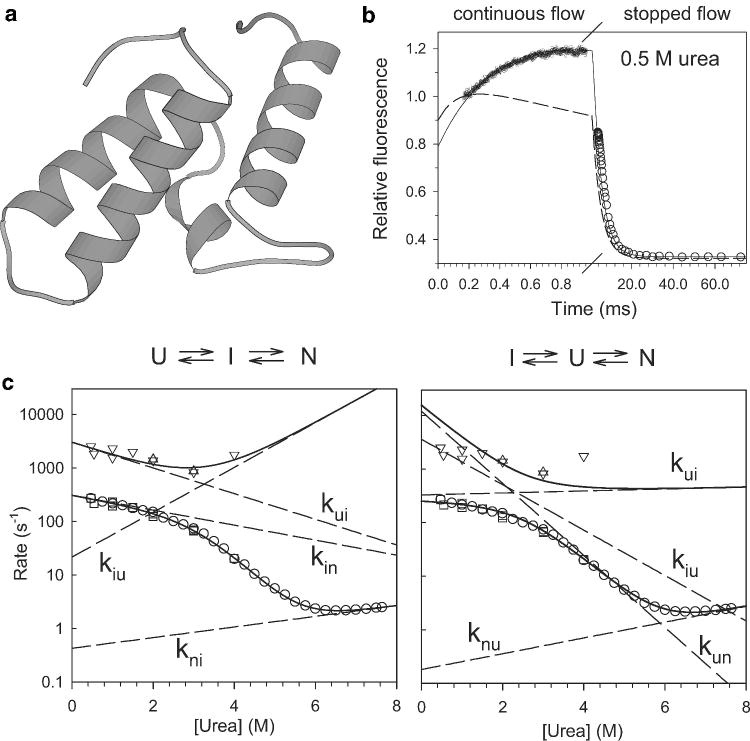
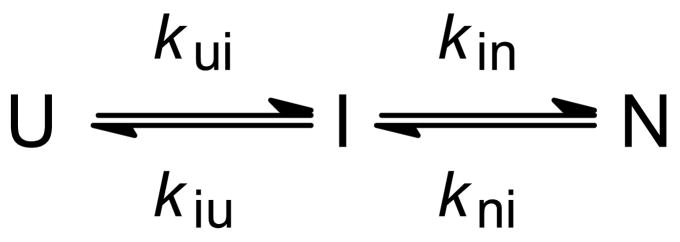
Kinetic mechanism of Im7 folding. a Ribbon diagram of Im7. b Representative kinetic trace measured by continuous-flow (●) and stopped-flow (○) fluorescence. The kinetics at this and all other urea concentrations measured is accurately predicted by on on-pathway mechanism (solid line) while schemes with off-pathway intermediates fail to reproduce the data (dashed line). c Observed (symbols) and predicted (solid lines) rates of folding and unfolding, based on mechanisms with on-pathway (left) and off-pathway (right) intermediates. Dashed lines indicate the corresponding elementary rate constants. Adapted from ref (Figures 2 and 3).
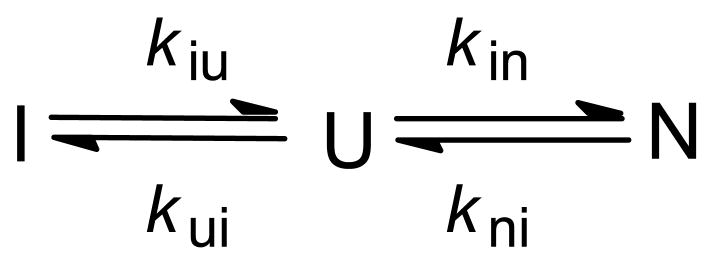
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
