

Allelic silencing at the tumor-suppressor locus 13q14.3 suggests an epigenetic tumor-suppressor mechanism
- PMID: 16684883
- PMCID: PMC1472515
- DOI: 10.1073/pnas.0600494103
Allelic silencing at the tumor-suppressor locus 13q14.3 suggests an epigenetic tumor-suppressor mechanism
Abstract
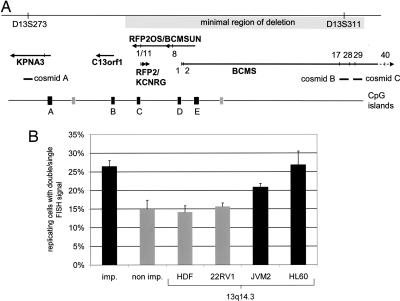
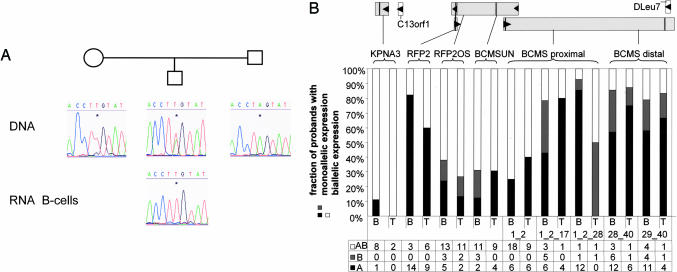
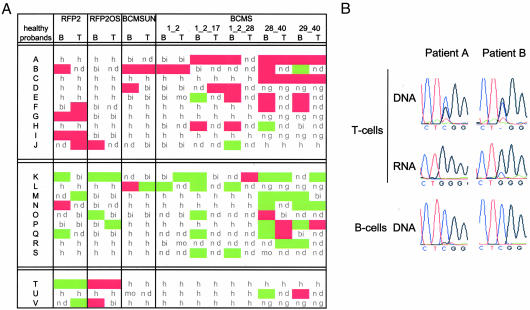
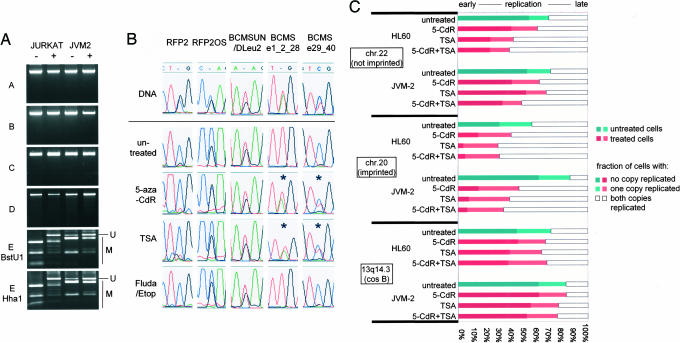
Genomic material from chromosome band 13q14.3 distal to the retinoblastoma locus is recurrently lost in a variety of human neoplasms, indicating an as-yet-unidentified tumor-suppressor mechanism. No pathogenic mutations have been found in the minimally deleted region until now. However, in B cell chronic lymphocytic leukemia tumors with loss of one copy of the critical region, respective candidate tumor-suppressor genes are down-regulated by a factor >2, which would be expected by a normal gene-dosage effect. This finding points to an epigenetic pathomechanism. We find that the two copies of the critical region replicate asynchronously, suggesting differential chromatin packaging of the two copies of 13q14.3. Although we also detect monoallelic silencing of genes localized in the critical region, monoallelic expression originates from either the maternal or paternal copy, excluding an imprinting mechanism. DNA methylation analyses revealed one CpG island of the region to be methylated. DNA demethylation of this CpG island and global histone hyperacetylation induced biallelic expression, whereas replication timing was not affected. We propose that differential replication timing represents an early epigenetic mark that distinguishes the two copies of 13q14.3, resulting in differential chromatin packaging and monoallelic expression. Accordingly, deletion of the single active copy of 13q14.3 results in significant down-regulation of the candidate genes and loss of function, providing a model for the interaction of genetic lesions and epigenetic silencing at 13q14.3 in B cell chronic lymphocytic leukemia.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- Stilgenbauer S., Nickolenko J., Wilhelm J., Wolf S., Weitz S., Döhner K., Boehm T., Döhner H., Lichter P. Oncogene. 1998;16:1891–1897. - PubMed
-
- Corcoran M. M., Hammarsund M., Zhu C., Lerner M., Kapanadze B., Wilson B., Larsson C., Forsberg L., Ibbotson R. E., Einhorn S., et al. Genes Chromosomes Cancer. 2004;40:285–297. - PubMed
-
- Liu Y., Corcoran M., Rasool O., Ivanova G., Ibbotson R., Grander D., Iyengar A., Baranova A., Kashuba V., Merup M., et al. Oncogene. 1997;15:2463–2473. - PubMed
-
- Migliazza A., Bosch F., Komatsu H., Cayanis E., Martinotti S., Toniato E., Guccione E., Qu X., Chien M., Murty V. V., et al. Blood. 2001;97:2098–2104. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
