

Kinetic and chemical analyses of the cytokinin dehydrogenase-catalysed reaction: correlations with the crystal structure
- PMID: 16686601
- PMCID: PMC1525011
- DOI: 10.1042/BJ20060280
Kinetic and chemical analyses of the cytokinin dehydrogenase-catalysed reaction: correlations with the crystal structure
Abstract
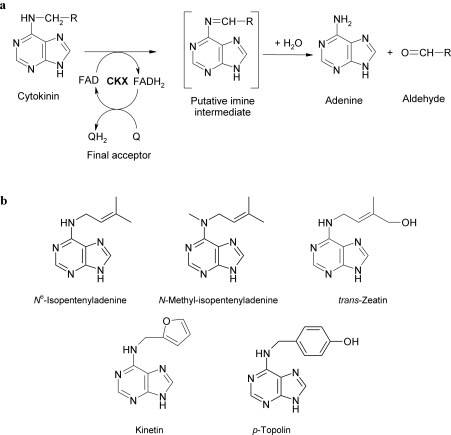
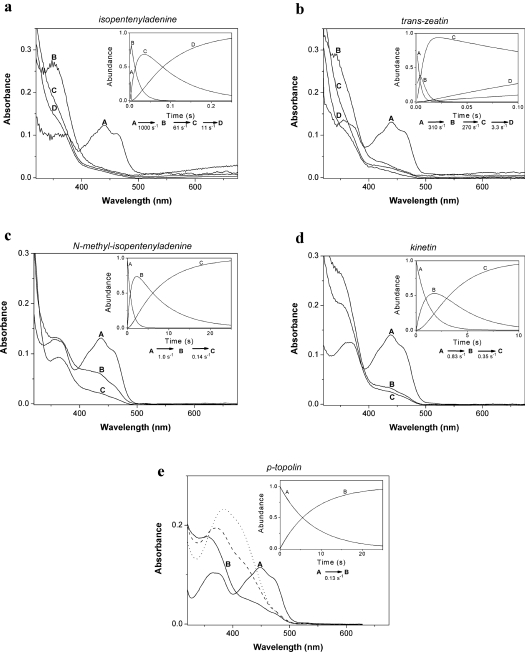
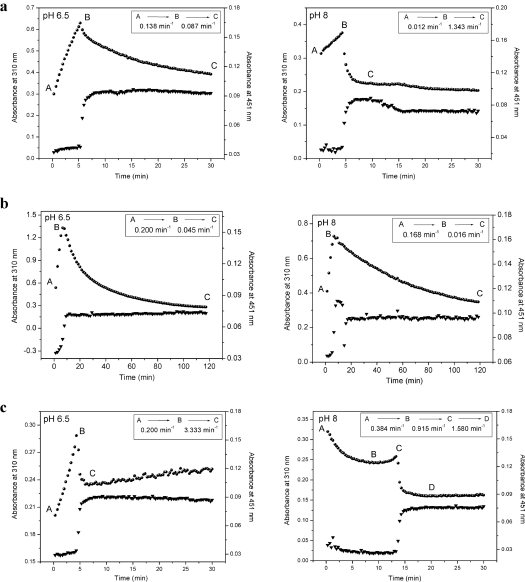
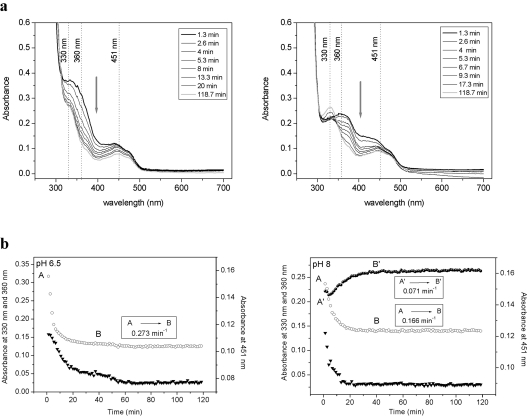
CKX (cytokinin dehydrogenase) is a flavoprotein that cleaves cytokinins to adenine and the corresponding side-chain aldehyde using a quinone-type electron acceptor. In the present study, reactions of maize (Zea mays) CKX with five different substrates (N6-isopentenyladenine, trans-zeatin, kinetin, p-topolin and N-methyl-isopentenyladenine) were studied. By using stopped-flow analysis of the reductive half-reaction, spectral intermediates were observed indicative of the transient formation of a binary enzyme-product complex between the cytokinin imine and the reduced enzyme. The reduction rate was high for isoprenoid cytokinins that showed formation of a charge-transfer complex of reduced enzyme with bound cytokinin imine. For the other cytokinins, flavin reduction was slow and no charge-transfer intermediates were observed. The binary complex of reduced enzyme and imine product intermediate decays relatively slowly to form an unbound product, cytokinin imine, which accumulates in the reaction mixture. The imine product only very slowly hydrolyses to adenine and an aldehyde derived from the cytokinin N6 side-chain. Mixing of the substrate-reduced enzyme with Cu2+/imidazole as an electron acceptor to monitor the oxidative half-reaction revealed a high rate of electron transfer for this type of electron acceptor when using N6-isopentenyladenine. The stability of the cytokinin imine products allowed their fragmentation analysis and structure assessment by Q-TOF (quadrupole-time-of-flight) MS/MS. Correlations of the kinetic data with the known crystal structure are discussed for reactions with different cytokinins.
Figures
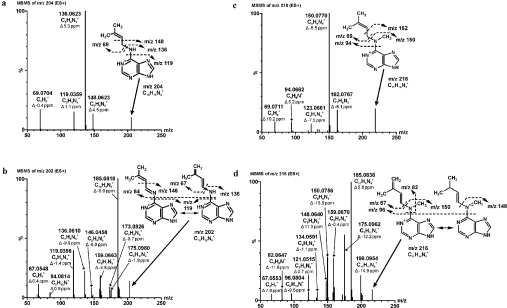
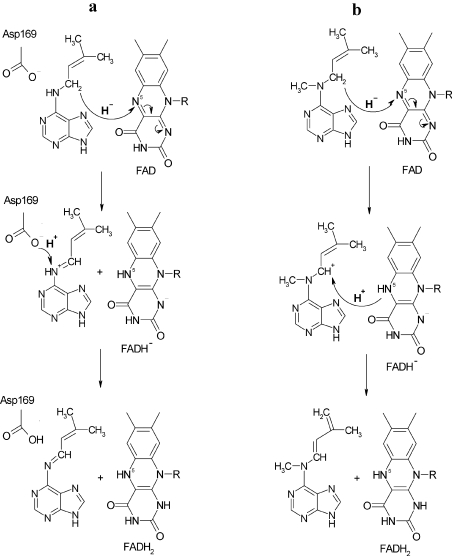
References
-
- van Berkel W. J. H., Benen J. A. E., Eppink M. H. M., Fraaije M. W. Flavoprotein kinetics. In: Chapman S. K., Reid G. A., editors. Methods in Molecular Biology, vol. 131, Flavoprotein Protocols. Totowa, NJ: Humana Press; 1999. pp. 61–86. - PubMed
-
- Hare P. D., van Staden J. Cytokinin oxidase: biochemical features and physiological significance. Physiol. Plant. 1994;91:128–136.
-
- Galuszka P., Frébort I., Šebela M., Peč P. Degradation of cytokinins by cytokinin oxidases in plants. Plant Growth Regul. 2000;32:315–327.
-
- Galuszka P., Frébort I., Šebela M., Sauer P., Jacobsen S., Peč P. Cytokinin oxidase or dehydrogenase?. Mechanism of cytokinin degradation in cereals. Eur. J. Biochem. 2001;268:450–461. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources