

Ranolazine improves abnormal repolarization and contraction in left ventricular myocytes of dogs with heart failure by inhibiting late sodium current
- PMID: 16686675
- PMCID: PMC1482456
- DOI: 10.1111/j.1540-8167.2006.00401.x
Ranolazine improves abnormal repolarization and contraction in left ventricular myocytes of dogs with heart failure by inhibiting late sodium current
Abstract
Background: Ventricular repolarization and contractile function are frequently abnormal in ventricular myocytes from human failing hearts as well as canine hearts with experimentally induced heart failure (HF). These abnormalities have been attributed to dysfunction involving various steps of the excitation-contraction coupling process, leading to impaired intracellular sodium and calcium homeostasis. We previously reported that the slow inactivating component of the Na(+) current (late I(Na)) is augmented in myocytes from failing hearts, and this appears to play a significant role in abnormal ventricular myocytes repolarization and function. We tested the effect of ranolazine, a novel drug being developed to treat angina, on (1) action potential duration (APD), (2) peak transient and late I(Na) (I(NaT) and I(NaL), respectively), (3) early afterdepolarizations (EADs), and (4) twitch contraction (TC), including after contractions and contracture.
Methods: Myocytes were isolated from the left ventricle of normal dogs and of dogs with chronic HF caused by multiple sequential intracoronary micro-embolizations. I(NaT) and I(NaL) were recorded using conventional whole-cell patch-clamp techniques. APs were recorded using the beta-escin perforated patch-clamp configuration at frequencies of 0.25 and 0.5 Hz. TCs were recorded using an edge movement detector at stimulation frequencies ranging from 0.5 to 2.0 Hz.
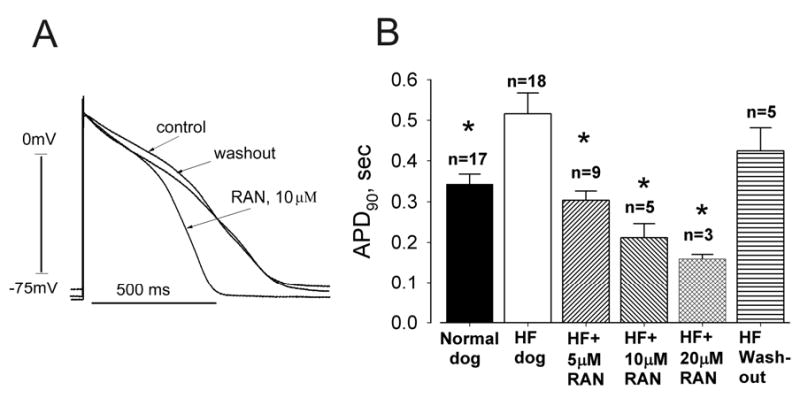
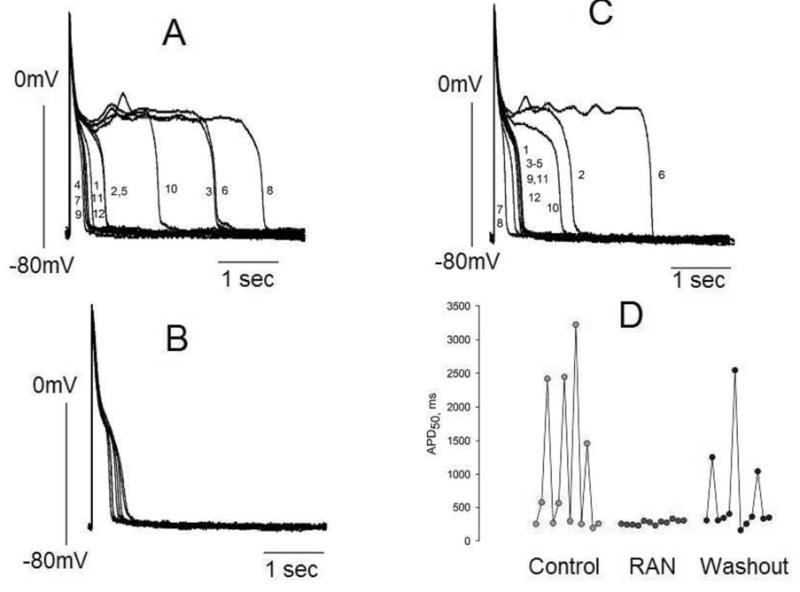
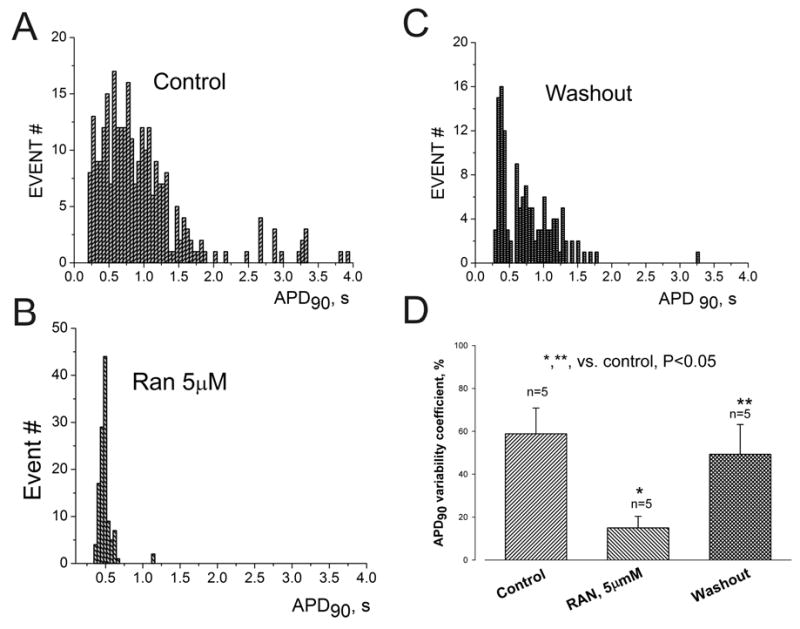
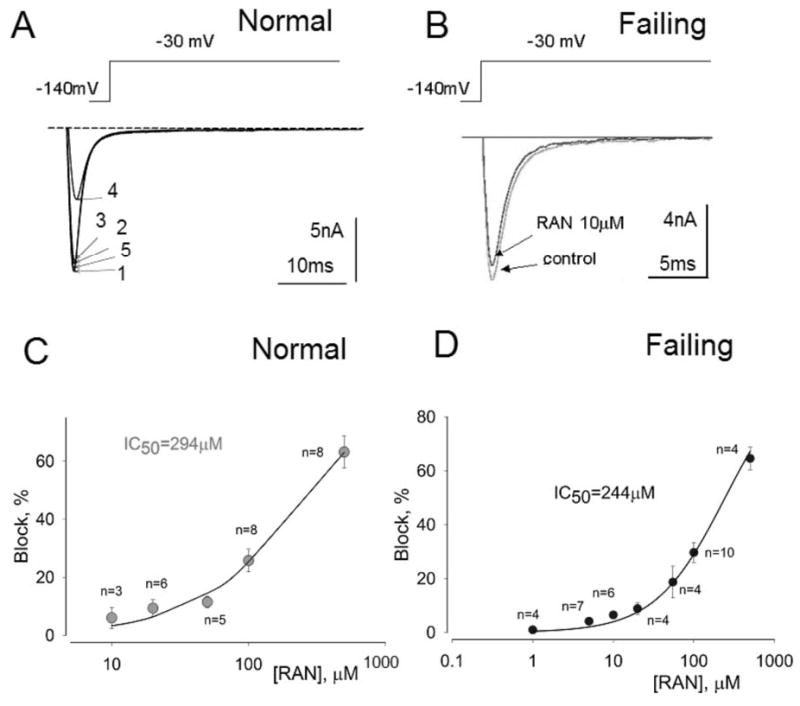
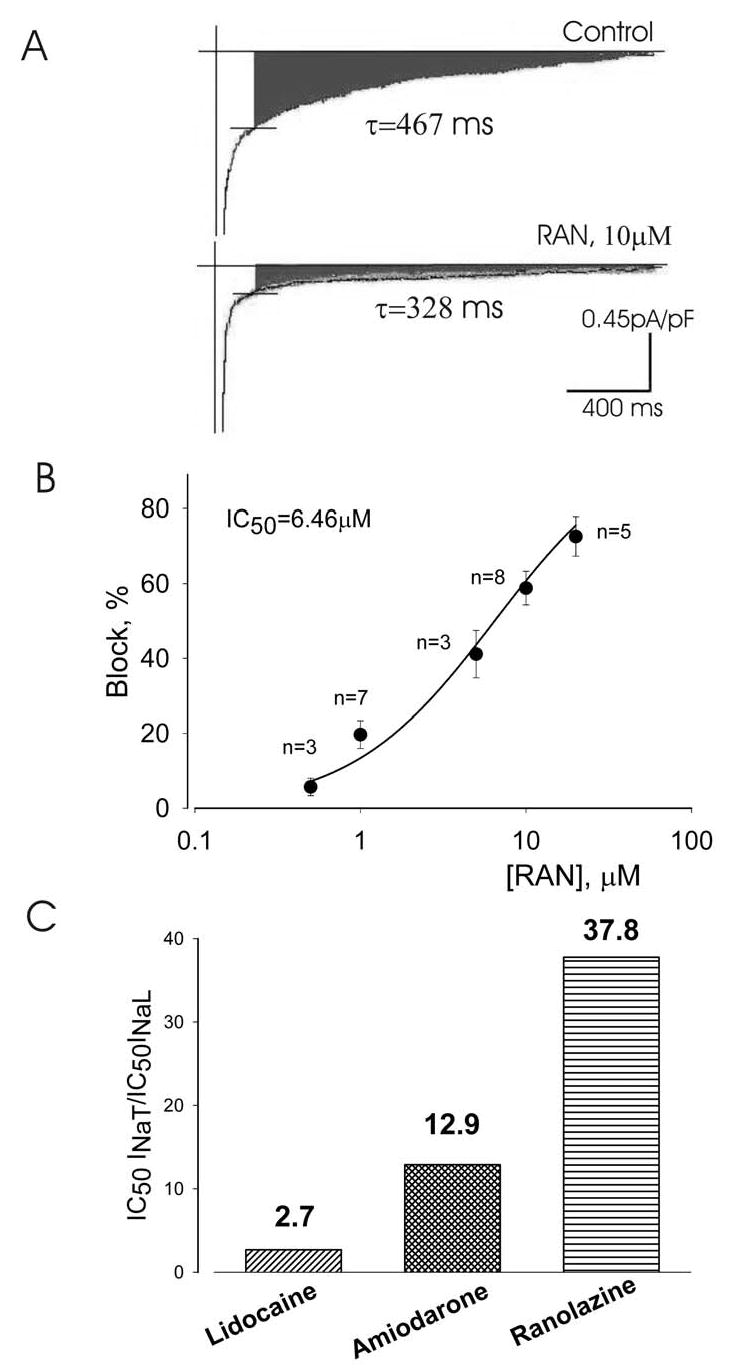
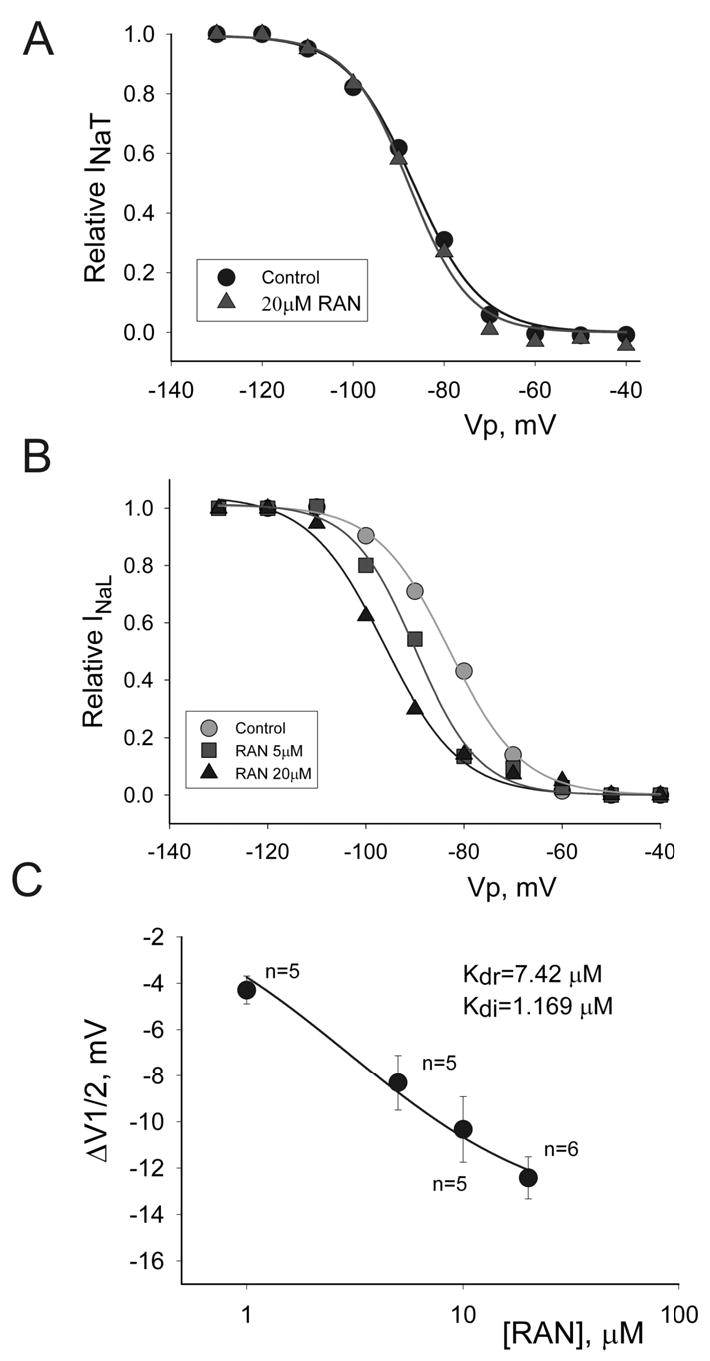
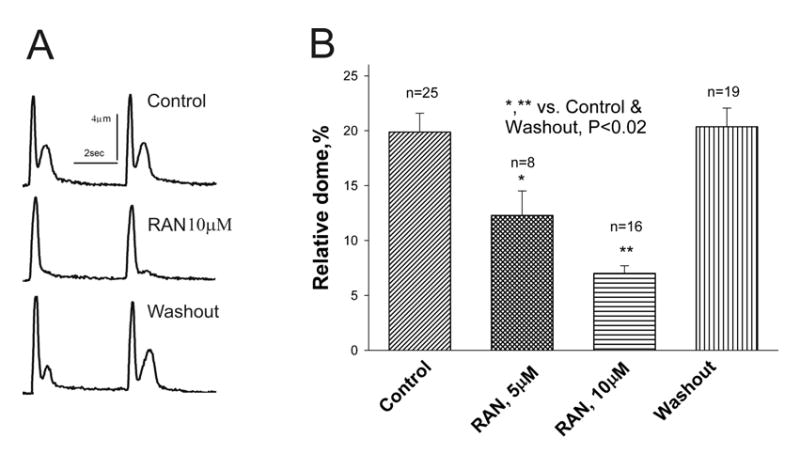
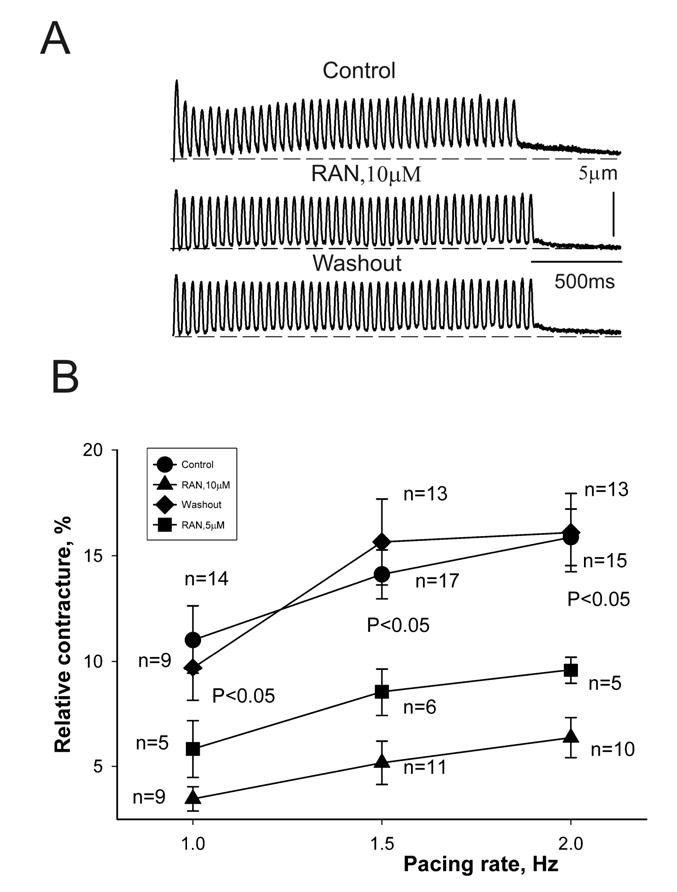
Results: Ranolazine significantly (P<0.05) and reversibly shortened the APD of myocytes stimulated at either 0.5 or 0.25 Hz in a concentration-dependent manner. At a stimulation frequency of 0.5 Hz, 5, 10, and 20 microM ranolazine shortened the APD(90) (APD measured at 90% repolarization) from 516+/-51 to 304+/-22, 212+/-34 and 160+/-11 ms, respectively, and markedly decreased beat-to-beat variability of APD(90), EADs, and dispersion of APDs. Ranolazine preferentially blocked I(NaL) relative to I(NaT) in a state-dependent manner, with a approximately 38-fold greater potency against I(NaL) to produce tonic block (IC(50)=6.5 microM) than I(NaT) (IC(50)=294 microM). When we evaluated inactivated state blockade of I(NaL) from the steady-state inactivation mid-potential shift using a theoretical model, ranolazine was found to bind more tightly to the inactivated state than the resting state of the sodium channel underlying I(NaL), with apparent dissociation constants K(dr)=7.47 microM and K(di)=1.71 microM, respectively. TCs of myocytes stimulated at 0.5 Hz were characterized by an initial spike followed by a dome-like after contraction, which was observed in 75% of myocytes from failing hearts and coincided with the long AP plateau and EADs. Ranolazine at 5 and 10 microM reversibly shortened the duration of TCs and abolished the after contraction. When the rate of myocyte stimulation was increased from 1.0 to 2.0 Hz, there was a progressive increase in diastolic "tension," that is, contracture. Ranolazine at 5 and 10 microM reversibly prevented this frequency-dependent contracture.
Figures

References
-
- Tomaselli GF, Zipes DP. What causes sudden death in heart failure? Circ Res. 2004;95:754–763. - PubMed
-
- Tomaselli GF, Marban E. Electrophysiological remodeling in hypertrophy and heart failure. Cardiovasc Res. 1999;42:270–283. - PubMed
-
- Galinier M, Vialette JC, Fourcade J, Cabrol P, Dongay B, Massabuau P, Boveda S, Doazan JP, Fauvel JM, Bounhoure JP. QT interval dispersion as a predictor of arrhythmic events in congestive heart failure. Importance of aetiology. Eur Heart J. 1998;19:1054–62. - PubMed
-
- Boccalandro F, Velasco A, Thomas C, Richards B, Radovancevic B. Relations among heart failure severity, left ventricular loading conditions, and repolarization length in advanced heart failure secondary to ischemic or idiopathic dilated cardiomyopathy. Am J Cardiol. 2003;92:544–7. - PubMed
-
- Beuckelmann DJ, Nabauer M, Erdmann E. Alterations of K+ currents in isolated human ventricular myocytes from patients with terminal heart failure. Circulation Research. 1993;73:379–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
