

Aurora-B/AIM-1 regulates the dynamic behavior of HP1alpha at the G2-M transition
- PMID: 16687578
- PMCID: PMC1483052
- DOI: 10.1091/mbc.e05-09-0906
Aurora-B/AIM-1 regulates the dynamic behavior of HP1alpha at the G2-M transition
Abstract
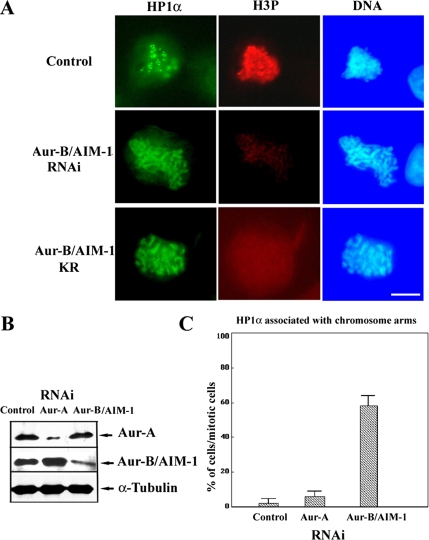
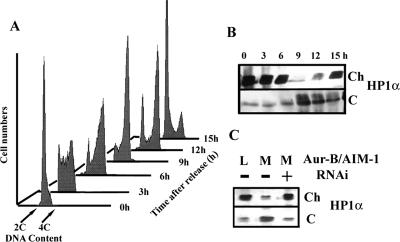
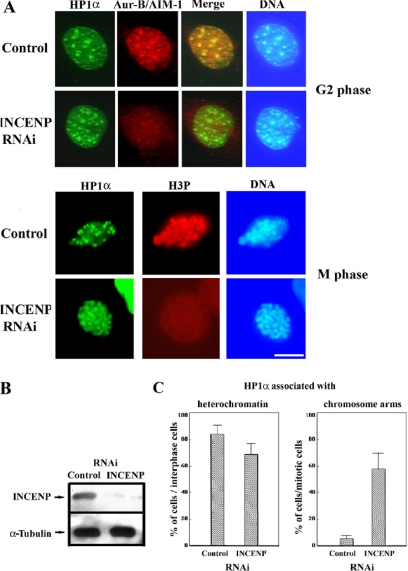
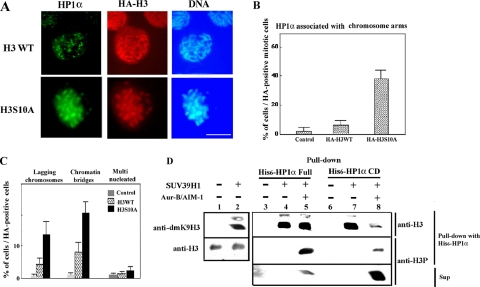
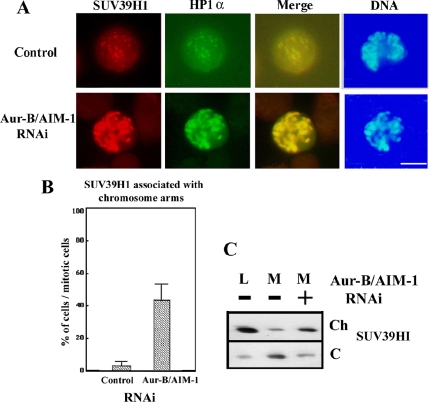
Heterochromatin protein 1 (HP1) plays an important role in heterochromatin formation and undergoes large-scale, progressive dissociation from heterochromatin in prophase cells. However, the mechanisms regulating the dynamic behavior of HP1 are poorly understood. In this study, the role of Aurora-B was investigated with respect to the dynamic behavior of HP1alpha. Mammalian Aurora-B, AIM-1, colocalizes with HP1alpha to the heterochromatin in G2. Depletion of Aurora-B/AIM-1 inhibited dissociation of HP1alpha from the chromosome arms at the G2-M transition. In addition, depletion of INCENP led to aberrant cellular localization of Aurora-B/AIM-1, but it did not affect heterochromatin targeting of HP1alpha. It was proposed in the binary switch hypothesis that phosphorylation of histone H3 at Ser-10 negatively regulates the binding of HP1alpha to the adjacent methylated Lys-9. However, Aurora-B/AIM-1-mediated phosphorylation of H3 induced dissociation of the HP1alpha chromodomain but not of the intact protein in vitro, indicating that the center and/or C-terminal domain of HP1alpha interferes with the effect of H3 phosphorylation on HP1alpha dissociation. Interestingly, Lys-9 methyltransferase SUV39H1 is abnormally localized together along the metaphase chromosome arms in Aurora-B/AIM-1-depleted cells. In conclusion, these results showed that Aurora-B/AIM-1 is necessary for regulated histone modifications involved in binding of HP1alpha by the N terminus of histone H3 during mitosis.
Figures
References
-
- Aagaard L., Schmid M., Warburton P., Jenuwein T. Mitotic phosphorylation of SUV39H1, a novel component of active centromeres, coincides with transient accumulation at mammalian centromeres. J. Cell Sci. 2000;113:817–829. - PubMed
-
- Adams R. R., Carmena M., Earnshaw W. C. Chromosomal passengers and the (aurora) ABCs of mitosis. Trends Cell Biol. 2001a;11:49–54. - PubMed
-
- Adams R. R., Wheatley S. P., Gouldsworthy A. M., Kandels-Lewis S. E., Carmena M., Smythe C., Gerloff D. L., Earnshaw W. C. INCENP binds the Aurora-related kinase AIRK2 and is required to target it to chromosomes, the central spindle and cleavage furrow. Curr. Biol. 2000;10:1075–1078. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
