

A mechanical explanation of RNA pseudoknot function in programmed ribosomal frameshifting
- PMID: 16688178
- PMCID: PMC7094908
- DOI: 10.1038/nature04735
A mechanical explanation of RNA pseudoknot function in programmed ribosomal frameshifting
Abstract
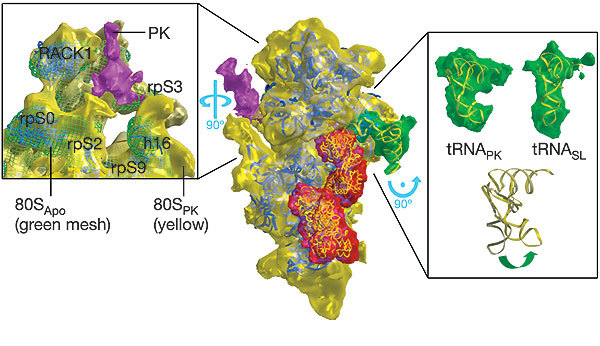
The triplet-based genetic code requires that translating ribosomes maintain the reading frame of a messenger RNA faithfully to ensure correct protein synthesis. However, in programmed -1 ribosomal frameshifting, a specific subversion of frame maintenance takes place, wherein the ribosome is forced to shift one nucleotide backwards into an overlapping reading frame and to translate an entirely new sequence of amino acids. This process is indispensable in the replication of numerous viral pathogens, including HIV and the coronavirus associated with severe acute respiratory syndrome, and is also exploited in the expression of several cellular genes. Frameshifting is promoted by an mRNA signal composed of two essential elements: a heptanucleotide 'slippery' sequence and an adjacent mRNA secondary structure, most often an mRNA pseudoknot. How these components operate together to manipulate the ribosome is unknown. Here we describe the observation of a ribosome-mRNA pseudoknot complex that is stalled in the process of -1 frameshifting. Cryoelectron microscopic imaging of purified mammalian 80S ribosomes from rabbit reticulocytes paused at a coronavirus pseudoknot reveals an intermediate of the frameshifting process. From this it can be seen how the pseudoknot interacts with the ribosome to block the mRNA entrance channel, compromising the translocation process and leading to a spring-like deformation of the P-site transfer RNA. In addition, we identify movements of the likely eukaryotic ribosomal helicase and confirm a direct interaction between the translocase eEF2 and the P-site tRNA. Together, the structural changes provide a mechanical explanation of how the pseudoknot manipulates the ribosome into a different reading frame.
Conflict of interest statement
Electron-density maps have been deposited in the European Bioinformatics Institute Electron Microscopy database, accession numbers EMD-1197, EMD-1198 and EMD-1199 (
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
