

Heavy metal stress and sulfate uptake in maize roots
- PMID: 16698905
- PMCID: PMC1489904
- DOI: 10.1104/pp.105.076240
Heavy metal stress and sulfate uptake in maize roots
Abstract
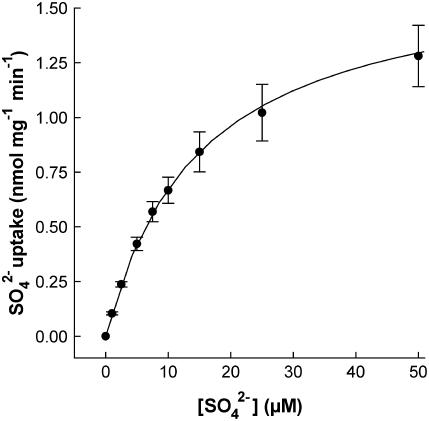
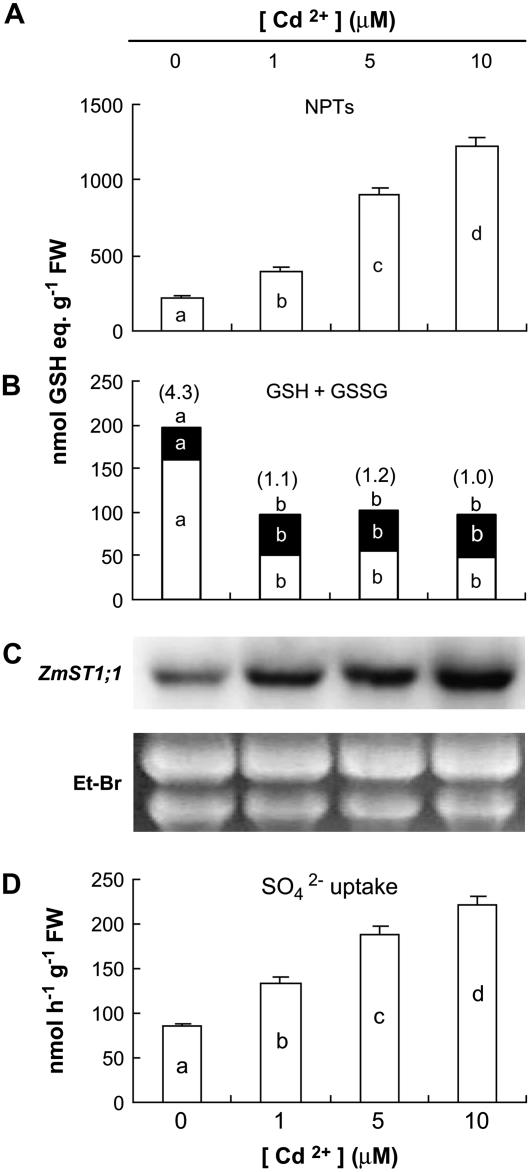
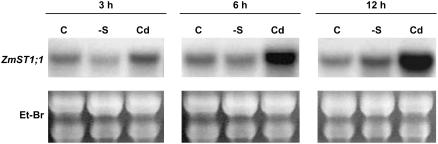
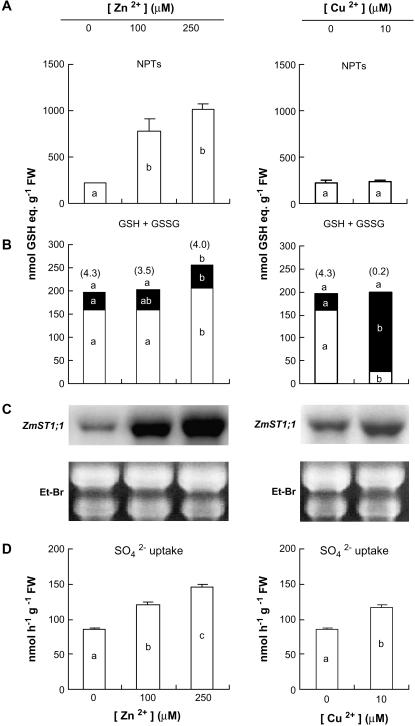
ZmST1;1, a putative high-affinity sulfate transporter gene expressed in maize (Zea mays) roots, was functionally characterized and its expression patterns were analyzed in roots of plants exposed to different heavy metals (Cd, Zn, and Cu) interfering with thiol metabolism. The ZmST1;1 cDNA was expressed in the yeast (Saccharomyces cerevisiae) sulfate transporter mutant CP154-7A. Kinetic analysis of sulfate uptake isotherm, determined on complemented yeast cells, revealed that ZmST1;1 has a high affinity for sulfate (Km value of 14.6 +/- 0.4 microm). Cd, Zn, and Cu exposure increased both ZmST1;1 expression and root sulfate uptake capacity. The metal-induced sulfate uptakes were accompanied by deep alterations in both thiol metabolism and levels of compounds such as reduced glutathione (GSH), probably involved as signals in sulfate uptake modulation. Cd and Zn exposure strongly increased the level of nonprotein thiols of the roots, indicating the induction of additional sinks for reduced sulfur, but differently affected root GSH contents that decreased or increased following Cd or Zn stress, respectively. Moreover, during Cd stress a clear relation between the ZmST1;1 mRNA abundance increment and the entity of the GSH decrement was impossible to evince. Conversely, Cu stress did not affect nonprotein thiol levels, but resulted in a deep contraction of GSH pools. Our data suggest that during heavy metal stress sulfate uptake by roots may be controlled by both GSH-dependent or -independent signaling pathways. Finally, some evidence suggesting that root sulfate availability in Cd-stressed plants may limit GSH biosynthesis and thus Cd tolerance are discussed.
Figures


Similar articles
-
Cadmium-induced sulfate uptake in maize roots.Plant Physiol. 2002 Aug;129(4):1872-9. doi: 10.1104/pp.002659. Plant Physiol. 2002. PMID: 12177501 Free PMC article.
-
Coordinated expression of sulfate uptake and components of the sulfate assimilatory pathway in maize.Plant Biol (Stuttg). 2004 Jul;6(4):408-14. doi: 10.1055/s-2004-820872. Plant Biol (Stuttg). 2004. PMID: 15248123
-
Chromate differentially affects the expression of a high-affinity sulfate transporter and isoforms of components of the sulfate assimilatory pathway in Zea mays (L.).Plant Biol (Stuttg). 2007 Sep;9(5):662-71. doi: 10.1055/s-2007-965440. Plant Biol (Stuttg). 2007. PMID: 17853366
-
Sulfur assimilation and glutathione metabolism under cadmium stress in yeast, protists and plants.FEMS Microbiol Rev. 2005 Sep;29(4):653-71. doi: 10.1016/j.femsre.2004.09.004. FEMS Microbiol Rev. 2005. PMID: 16102596 Review.
-
Plant responses to abiotic stresses: heavy metal-induced oxidative stress and protection by mycorrhization.J Exp Bot. 2002 May;53(372):1351-65. J Exp Bot. 2002. PMID: 11997381 Review.
Cited by
-
The dynamics of DNA methylation in maize roots under Pb stress.Int J Mol Sci. 2014 Dec 17;15(12):23537-54. doi: 10.3390/ijms151223537. Int J Mol Sci. 2014. PMID: 25526567 Free PMC article.
-
Influence of sulfur and cadmium on antioxidants, phytochelatins and growth in Indian mustard.AoB Plants. 2015 Jan 12;7:plv001. doi: 10.1093/aobpla/plv001. AoB Plants. 2015. PMID: 25587194 Free PMC article.
-
Sulfur Protects Pakchoi (Brassica chinensis L.) Seedlings against Cadmium Stress by Regulating Ascorbate-Glutathione Metabolism.Int J Mol Sci. 2017 Jul 26;18(8):1628. doi: 10.3390/ijms18081628. Int J Mol Sci. 2017. PMID: 28933771 Free PMC article.
-
Cross-talk between sulfur assimilation and ethylene signaling in plants.Plant Signal Behav. 2013 Jan;8(1):e22478. doi: 10.4161/psb.22478. Epub 2012 Oct 26. Plant Signal Behav. 2013. PMID: 23104111 Free PMC article. Review.
-
Genome-wide identification and function analysis of HMAD gene family in cotton (Gossypium spp.).BMC Plant Biol. 2021 Aug 20;21(1):386. doi: 10.1186/s12870-021-03170-8. BMC Plant Biol. 2021. PMID: 34416873 Free PMC article.
References
-
- Bolchi A, Petrucco S, Tenca PL, Foroni C, Ottonello S (1999) Coordinate modulation of maize sulphate permease and ATP sulfurylase mRNAs in response to variations in sulfur nutritional status: stereospecific down-regulation by L-cysteine. Plant Mol Biol 39: 527–537 - PubMed
-
- Clemens S (2001) Molecular mechanisms of plant metal tolerance and homeostasis. Planta 212: 475–486 - PubMed
-
- Gallego SM, Benavides MP, Tomaro ML (1996) Effect of heavy metal ion excess on sunflower leaves: evidence for involvement of oxidative stress. Plant Sci 121: 151–159
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
