

Runaway domain swapping in amyloid-like fibrils of T7 endonuclease I
- PMID: 16698921
- PMCID: PMC1472426
- DOI: 10.1073/pnas.0602607103
Runaway domain swapping in amyloid-like fibrils of T7 endonuclease I
Abstract
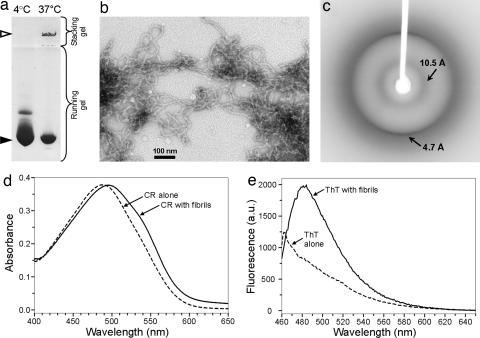
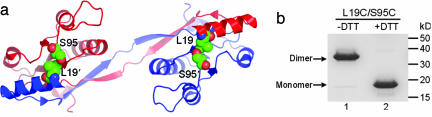
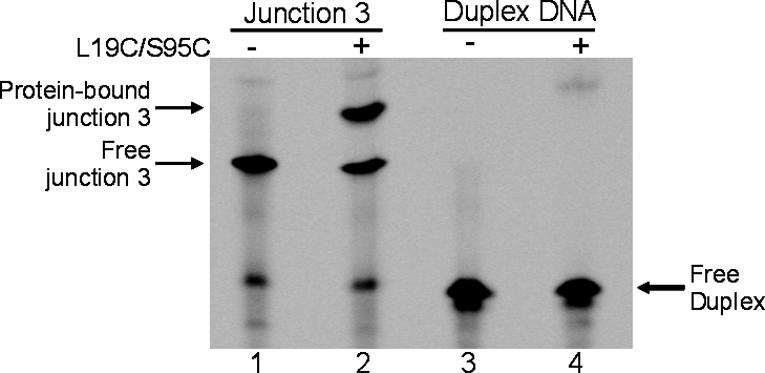
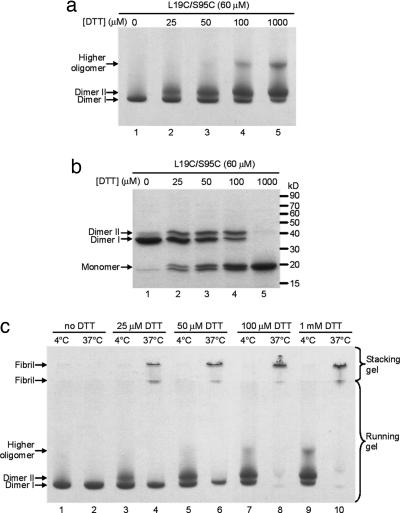
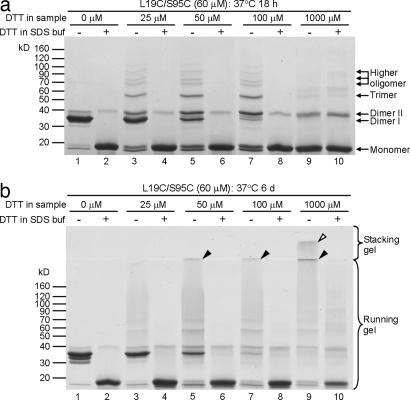
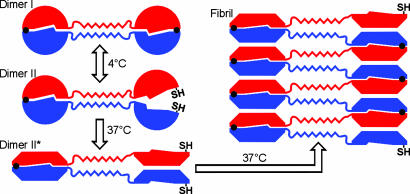
Amyloid fibrils are associated with >20 fatal human disorders, including Alzheimer's, Parkinson's, and prion diseases. Knowledge of how soluble proteins assemble into amyloid fibrils remains elusive despite its potential usefulness for developing diagnostics and therapeutics. In at least some fibrils, runaway domain swapping has been proposed as a possible mechanism for fibril formation. In runaway domain swapping, each protein molecule swaps a domain into the complementary domain of the adjacent molecule along the fibril. Here we show that T7 endonuclease I, a naturally domain-swapped dimeric protein, can form amyloid-like fibrils. Using protein engineering, we designed a double-cysteine mutant that forms amyloid-like fibrils in which molecules of T7 endonuclease I are linked by intermolecular disulfide bonds. Because the disulfide bonds are designed to form only at the domain-swapped dimer interface, the resulting covalently linked fibrils show that T7 endonuclease I forms fibrils by a runaway domain swap. In addition, we show that the disulfide mutant exists in two conformations, only one of which is able to form fibrils. We also find that domain-swapped dimers, if locked in a close-ended dimeric form, are unable to form fibrils. Our study provides strong evidence for runaway domain swapping in the formation of an amyloid-like fibril and, consequently, a molecular explanation for specificity and stability of fibrils. In addition, our results suggest that inhibition of fibril formation for domain-swapped proteins may be achieved by stabilizing domain-swapped dimers.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures

Similar articles
-
The GB1 amyloid fibril: recruitment of the peripheral beta-strands of the domain swapped dimer into the polymeric interface.J Mol Biol. 2005 May 6;348(3):687-98. doi: 10.1016/j.jmb.2005.02.071. J Mol Biol. 2005. PMID: 15826664
-
Amyloid-like fibrils from a domain-swapping protein feature a parallel, in-register conformation without native-like interactions.J Biol Chem. 2011 Aug 19;286(33):28988-28995. doi: 10.1074/jbc.M111.261750. Epub 2011 Jun 28. J Biol Chem. 2011. PMID: 21715337 Free PMC article.
-
Limited Proteolysis Reveals That Amyloids from the 3D Domain-Swapping Cystatin B Have a Non-Native β-Sheet Topology.J Mol Biol. 2015 Jul 31;427(15):2418-2434. doi: 10.1016/j.jmb.2015.05.014. Epub 2015 May 22. J Mol Biol. 2015. PMID: 26004542
-
3D domain swapping, protein oligomerization, and amyloid formation.Acta Biochim Pol. 2001;48(4):807-27. Acta Biochim Pol. 2001. PMID: 11995994 Review.
-
3D domain swapping: as domains continue to swap.Protein Sci. 2002 Jun;11(6):1285-99. doi: 10.1110/ps.0201402. Protein Sci. 2002. PMID: 12021428 Free PMC article. Review.
Cited by
-
Serpins show structural basis for oligomer toxicity and amyloid ubiquity.FEBS Lett. 2008 Jul 23;582(17):2537-41. doi: 10.1016/j.febslet.2008.06.021. Epub 2008 Jun 23. FEBS Lett. 2008. PMID: 18573252 Free PMC article.
-
Protein folding and misfolding on surfaces.Int J Mol Sci. 2008 Dec;9(12):2515-2542. doi: 10.3390/ijms9122515. Epub 2008 Dec 9. Int J Mol Sci. 2008. PMID: 19330090 Free PMC article.
-
Molecular determinants and genetic modifiers of aggregation and toxicity for the ALS disease protein FUS/TLS.PLoS Biol. 2011 Apr;9(4):e1000614. doi: 10.1371/journal.pbio.1000614. Epub 2011 Apr 26. PLoS Biol. 2011. PMID: 21541367 Free PMC article.
-
The impact of solubility and electrostatics on fibril formation by the H3 and H4 histones.Protein Sci. 2011 Dec;20(12):2060-73. doi: 10.1002/pro.743. Epub 2011 Nov 9. Protein Sci. 2011. PMID: 21953551 Free PMC article.
-
Ribonuclease A suggests how proteins self-chaperone against amyloid fiber formation.Protein Sci. 2012 Jan;21(1):26-37. doi: 10.1002/pro.754. Epub 2011 Nov 23. Protein Sci. 2012. PMID: 22095666 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
