

Recombinant adenoviral vectors can induce expression of p73 via the E4-orf6/7 protein
- PMID: 16699015
- PMCID: PMC1472169
- DOI: 10.1128/JVI.02016-05
Recombinant adenoviral vectors can induce expression of p73 via the E4-orf6/7 protein
Abstract
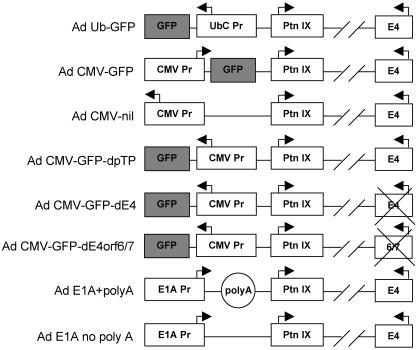
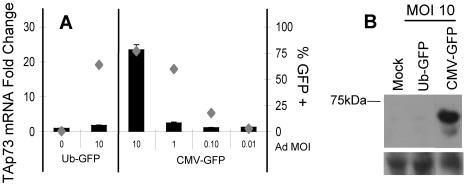
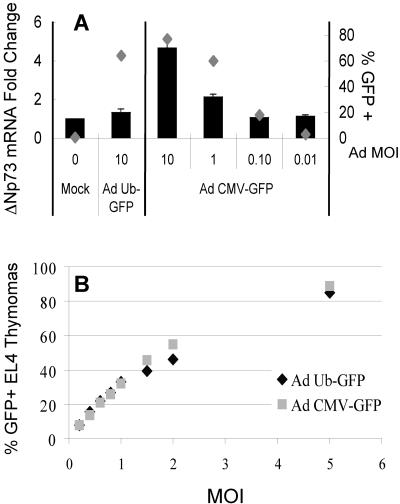
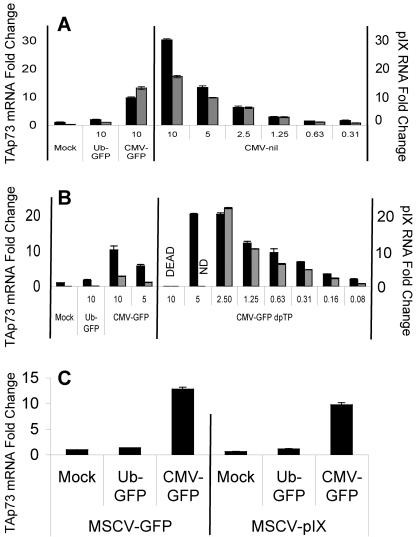
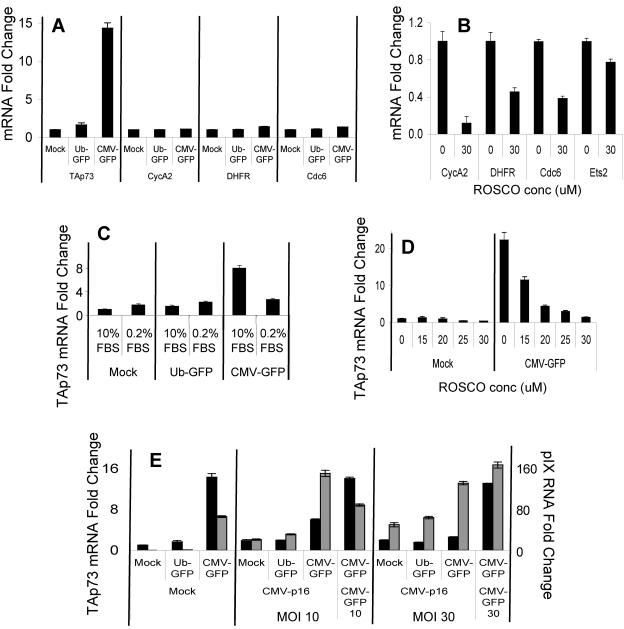
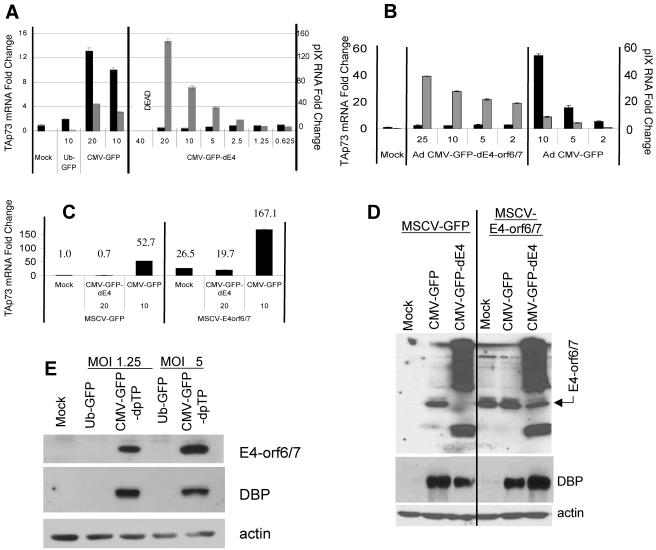
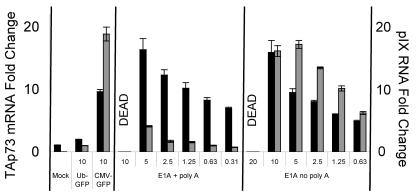
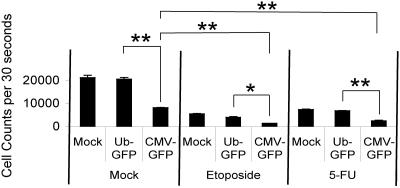
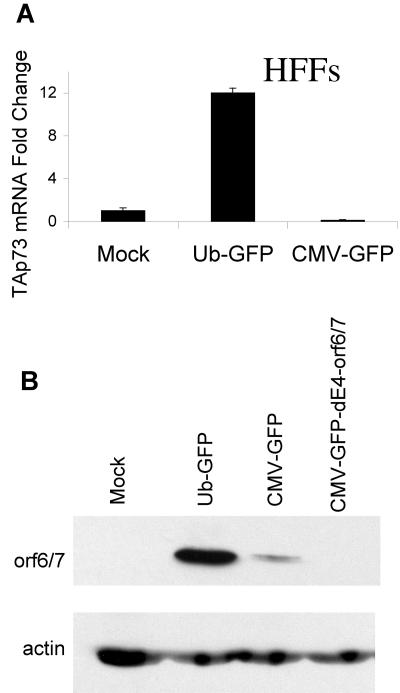
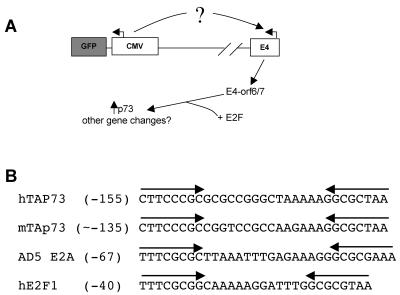
Despite the utility of recombinant adenoviral vectors in basic research, their therapeutic promise remains unfulfilled. Most engineered adenoviral vectors use a heterologous promoter to transcribe a foreign gene. We show that adenoviruses containing the cytomegalovirus immediate-early promoter induce the expression of the proapoptotic cellular protein TAp73 via the cyclin-dependent kinase-retinoblastoma protein-E2F pathway in murine embryonic fibroblasts. Cells transduced with these vectors also expressed high levels of the adenoviral E4-orf6/7 and E2A proteins. By contrast, adenoviruses containing the ubiquitin C promoter failed to elicit these effects. E4-orf6/7 is necessary and sufficient for increased TAp73 expression, as shown by using retrovirus-mediated E4-orf6/7 expression and adenovirus with the E4-orf6/7 gene deleted. Activation of TAp73 likely occurs via E4-orf6/7-induced dimerization of E2F and subsequent binding to the inverted E2F-responsive elements within the TAp73 promoter. In addition, adenoviral vectors containing the cytomegalovirus immediate-early promoter, but not the ubiquitin C promoter, cooperated with chemotherapeutic agents to decrease cellularity in vitro. In contrast to murine embryonic fibroblasts, adenoviruses containing the ubiquitin C promoter, but not the cytomegalovirus immediate-early promoter, induced both E4-orf6/7 and TAp73 in human foreskin fibroblasts, emphasizing the importance of cellular context for promoter-dependent effects. Because TAp73 is important for the efficacy of chemotherapy, adenoviruses that increase TAp73 expression may enhance cancer therapies by promoting apoptosis. However, such adenoviruses may impair the long-term survival of transduced cells during gene replacement therapies. Our findings reveal previously unknown effects of foreign promoters in recombinant adenoviral vectors and suggest means to improve the utility of engineered adenoviruses by better controlling their impact on viral and cellular gene expression.
Figures
References
-
- Ben-Israel, H., and T. Kleinberger. 2002. Adenovirus and cell cycle control. Front. Biosci. 7:d1369-d1395. - PubMed
-
- Bergamaschi, D., M. Gasco, L. Hiller, A. Sullivan, N. Syed, G. Trigiante, I. Yulug, M. Merlano, G. Numico, A. Comino, M. Attard, O. Reelfs, B. Gusterson, A. K. Bell, V. Heath, M. Tavassoli, P. J. Farrell, P. Smith, X. Lu, and T. Crook. 2003. p53 polymorphism influences response in cancer chemotherapy via modulation of p73-dependent apoptosis. Cancer Cell 3:387-402. - PubMed
-
- Boyer, J., K. Rohleder, and G. Ketner. 1999. Adenovirus E4 34k and E4 11k inhibit double strand break repair and are physically associated with the cellular DNA-dependent protein kinase. Virology 263:307-312. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
