

Roles of human parainfluenza virus type 3 bases 13 to 78 in replication and transcription: identification of an additional replication promoter element and evidence for internal transcription initiation
- PMID: 16699019
- PMCID: PMC1472146
- DOI: 10.1128/JVI.00204-06
Roles of human parainfluenza virus type 3 bases 13 to 78 in replication and transcription: identification of an additional replication promoter element and evidence for internal transcription initiation
Abstract
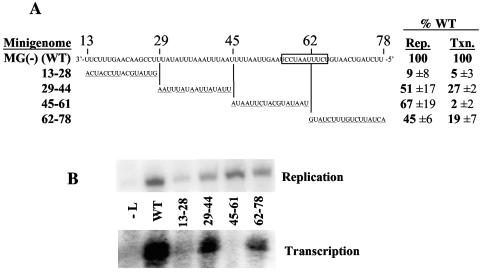
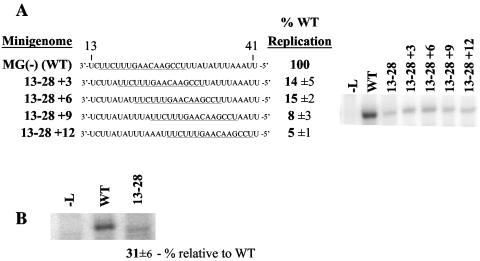
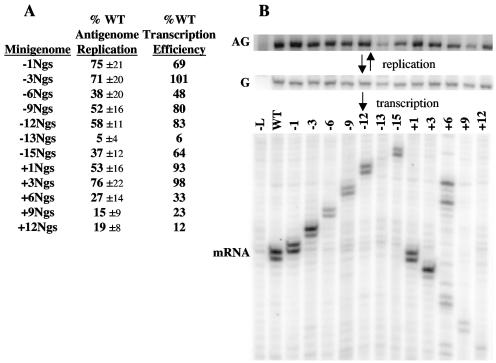
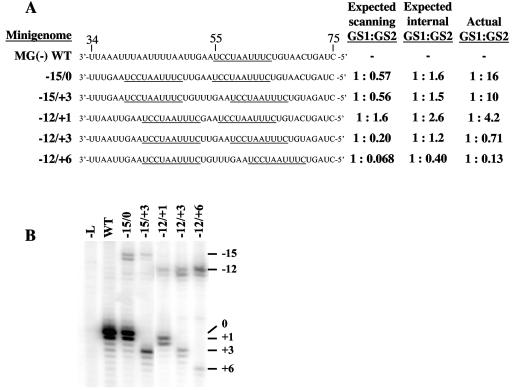
The genomic promoter of human parainfluenza virus type 3 (HPIV3) contains multiple cis-elements controlling transcription and replication. Previous work showed that regions 1 to 12 and 79 to 96 were critical in promoting replication of an HPIV3 minireplicon, while the intergenic sequence and N gene start signal (IS/Ngs, bases 49 to 61) were important for transcription. Because these data were collected primarily using point mutations, not every base from position 1 to 96 was analyzed, and some important control elements may have been missed. To clarify the role of bases 13 to 78 in transcription and replication, a series of mutations were made which collectively scanned this entire region. Mutation of bases 13 to 28 resulted in markedly decreased HPIV3 minireplicon replication, indicating these bases constitute an additional cis-element involved in the synthesis of the HPIV3 antigenomic RNA. The position dependence of the IS/Ngs was also examined. Analysis of mutants in which the IS/Ngs was shifted 5' or 3' showed that this segment could be moved without significantly disrupting transcription initiation. Additional mutants which contained two successive IS/Ngs segments were created to test whether the polymerase accessed the gene start signal by proceeding along the template 3' to 5' or by binding internally at the gene start signal. Based on analysis of the double gene start mutants, we propose a model of internal transcription initiation in which the polymerase enters the template at approximately the location of the natural N gene start but then scans the template bidirectionally to find a gene start signal and initiate transcription.
Figures

References
-
- Blumberg, B. M., C. Giorgi, and D. Kolakofsky. 1983. N protein of vesicular stomatitis virus selectively encapsidates leader RNA in vitro. Cell 32:559-567. - PubMed
-
- Calain, P., and L. Roux. 1995. Functional characterisation of the genomic and antigenomic promoters of Sendai virus. Virology 212:163-173. - PubMed
-
- Chanock, R. M., B. R. Murphy, and P. L. Collins. 2001. Parainfluenza viruses, p. 1341-1380. In D. M. Knipe, P. M. Howley, D. E. Griffin, R. A. Lamb, M. A. Martin, B. Roisman, and S. E. Straus (ed.), Fields virology, 4th ed., vol. 1. Lippincott Williams & Wilkins, Philadelphia, Pa.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
