

Palmitoylation of huntingtin by HIP14 is essential for its trafficking and function
- PMID: 16699508
- PMCID: PMC2279235
- DOI: 10.1038/nn1702
Palmitoylation of huntingtin by HIP14 is essential for its trafficking and function
Abstract
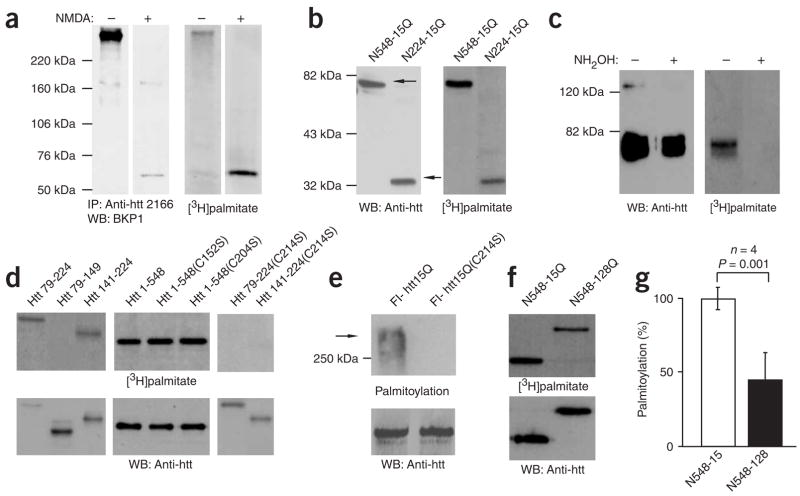
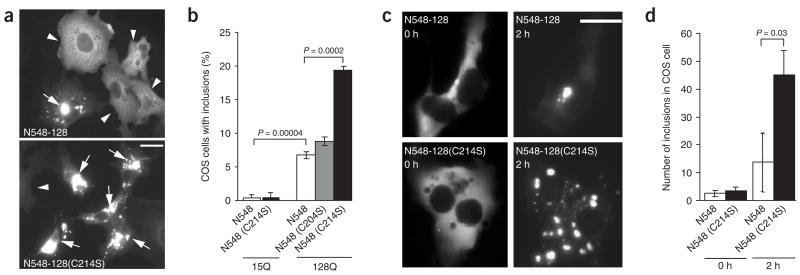
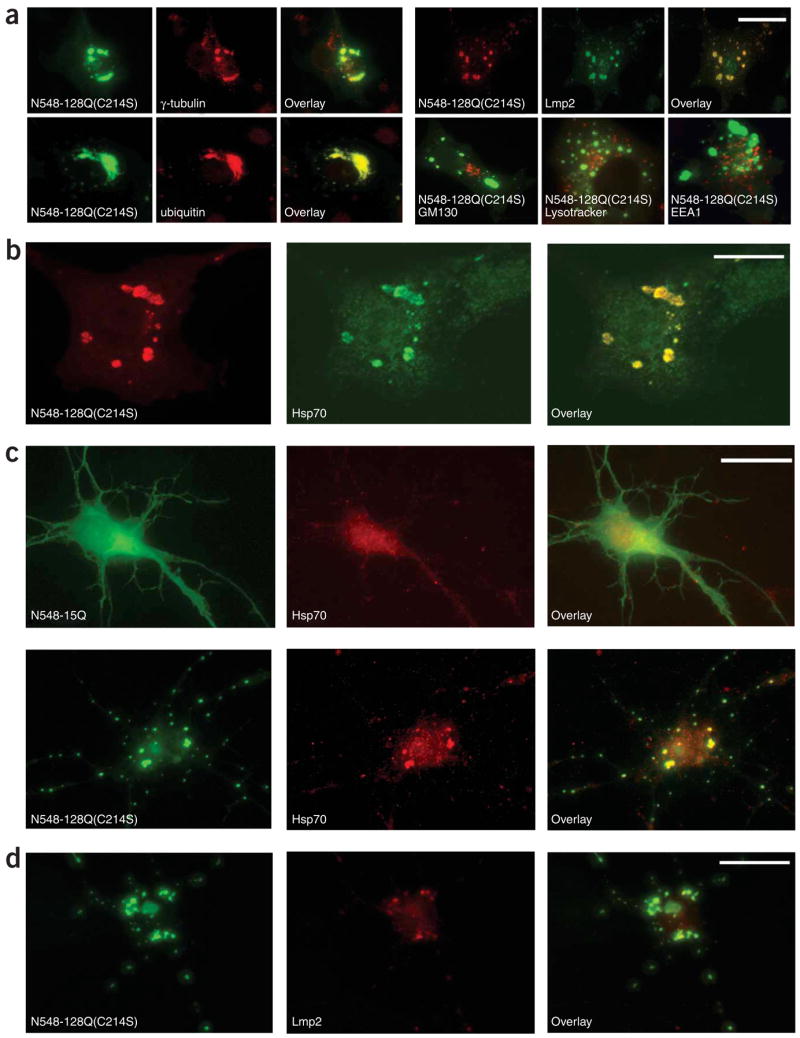
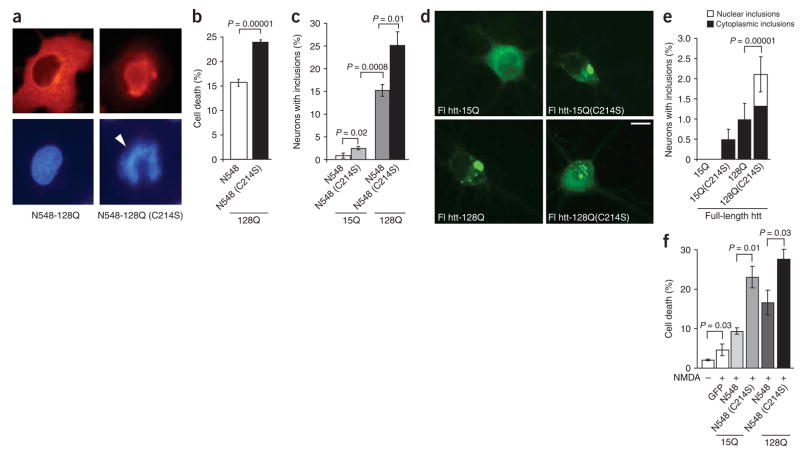
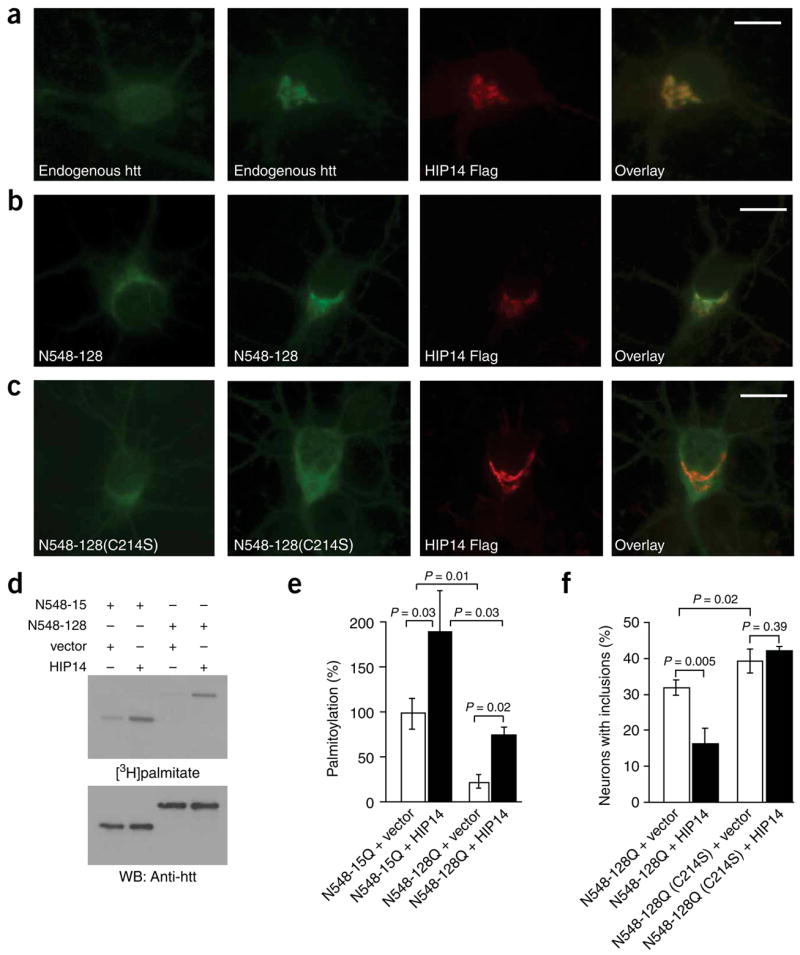
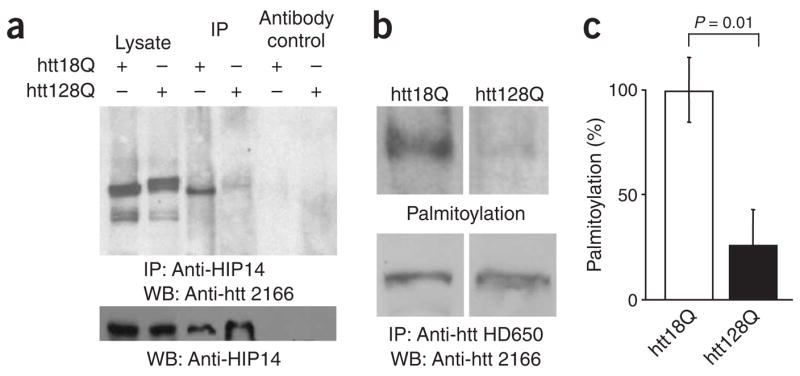
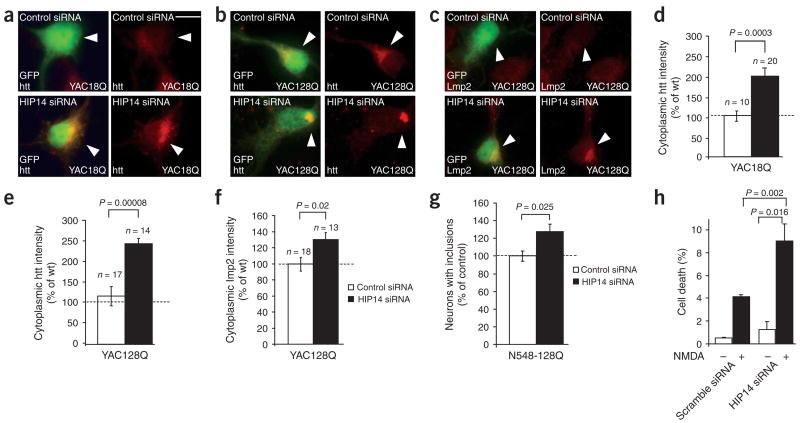
Post-translational modification by the lipid palmitate is crucial for the correct targeting and function of many proteins. Here we show that huntingtin (htt) is normally palmitoylated at cysteine 214, which is essential for its trafficking and function. The palmitoylation and distribution of htt are regulated by the palmitoyl transferase huntingtin interacting protein 14 (HIP14). Expansion of the polyglutamine tract of htt, which causes Huntington disease, results in reduced interaction between mutant htt and HIP14 and consequently in a marked reduction in palmitoylation. Mutation of the palmitoylation site of htt, making it palmitoylation resistant, accelerates inclusion formation and increases neuronal toxicity. Downregulation of HIP14 in mouse neurons expressing wild-type and mutant htt increases inclusion formation, whereas overexpression of HIP14 substantially reduces inclusions. These results suggest that the expansion of the polyglutamine tract in htt results in decreased palmitoylation, which contributes to the formation of inclusion bodies and enhanced neuronal toxicity.
Figures
References
-
- El-Husseini A, Bredt DS. Protein palmitoylation: a regulator of neuronal development and function. Nat Rev Neurosci. 2002;3:791–802. - PubMed
-
- Huang K, El-Husseini A. Modulation of neuronal protein trafficking and function by palmitoylation. Curr Opin Neurobiol. 2005;15:527–535. - PubMed
-
- Huang K, et al. Huntingtin-interacting protein HIP14 is a palmitoyl transferase involved in palmitoylation and trafficking of multiple neuronal proteins. Neuron. 2004;44:977–986. - PubMed
-
- The Huntington’s Disease Collaborative Research Group. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell. 1993;72:971–983. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
