

Ca2+-activated Cl- current from human bestrophin-4 in excised membrane patches
- PMID: 16702355
- PMCID: PMC2151534
- DOI: 10.1085/jgp.200609527
Ca2+-activated Cl- current from human bestrophin-4 in excised membrane patches
Abstract
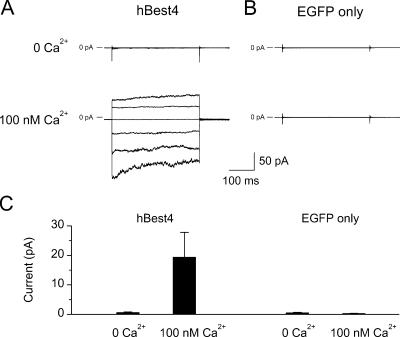
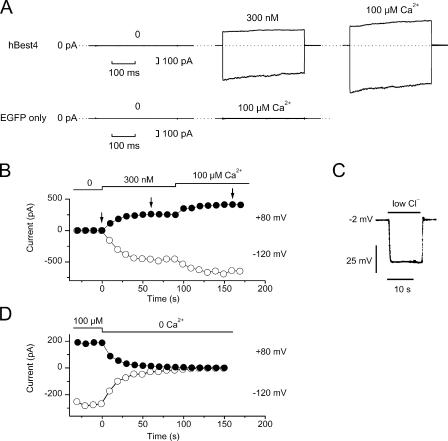
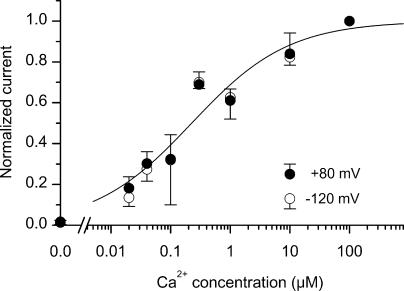
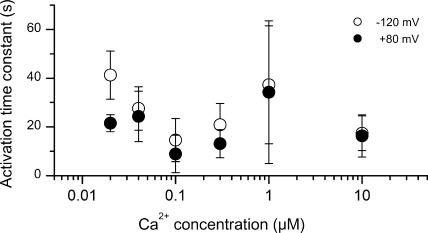
Bestrophins are a newly discovered family of Cl(-) channels, some members of which are activated by intracellular Ca(2+). So far, all studies were carried out with whole-cell recordings from plasmid-transfected cultured cells, so it is unclear whether Ca(2+) activates bestrophin through a metabolic mechanism or in a more direct way. We report here experiments that addressed this question with excised, inside-out membrane patches. We chose human bestrophin-4 (hBest4) for heterologous expression because it gave particularly large Cl(-) currents when expressed, thus allowing detection even in excised membrane patches. hBest4 gave a negligible Cl(-) current in a Ca(2+)-free solution on the cytoplasmic (bath) side, but produced a Cl(-) current that was activated by Ca(2+) in a dose-dependent manner, with a K(1/2) of 230 nM. Thus, Ca(2+) appears to activate the bestrophin Cl(-) channel without going through a freely diffusible messenger or through protein phosphorylation. Because the activation and deactivation kinetics were very slow, however, we cannot exclude the involvement of a membrane-associated messenger.
Figures
References
-
- Clapham, D.E. 2003. TRP channels as cellular sensors. Nature. 426:517–524. - PubMed
-
- Deutman, A.F. 1969. Electro-oculography in families with vitelliform dystrophy of the fovea. Detection of the carrier state. Arch. Ophthalmol. 81:305–316. - PubMed
-
- Francois, J., A. De Rouck, and D. Fernandez-Sasso. 1967. Electro-oculography in vitelliform degeneration of the macula. Arch. Ophthalmol. 77:726–733. - PubMed
-
- Gallemore, R.P., B.A. Hughes, and S.S. Miller. 1998. a. Light-induced responses of the retinal pigment epithelium. In The Retinal Pigment Epithelium. M.F. Marmor and T.J. Wolfensberger, editors. Oxford University Press, Oxford. 175–198.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
