

The Evf-2 noncoding RNA is transcribed from the Dlx-5/6 ultraconserved region and functions as a Dlx-2 transcriptional coactivator
- PMID: 16705037
- PMCID: PMC1475760
- DOI: 10.1101/gad.1416106
The Evf-2 noncoding RNA is transcribed from the Dlx-5/6 ultraconserved region and functions as a Dlx-2 transcriptional coactivator
Abstract
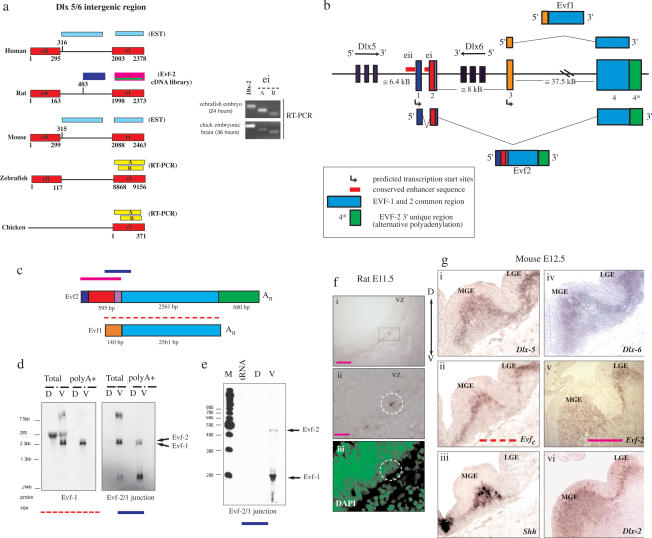
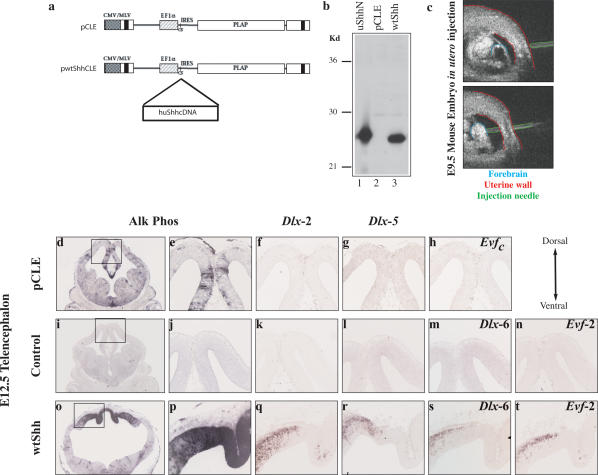
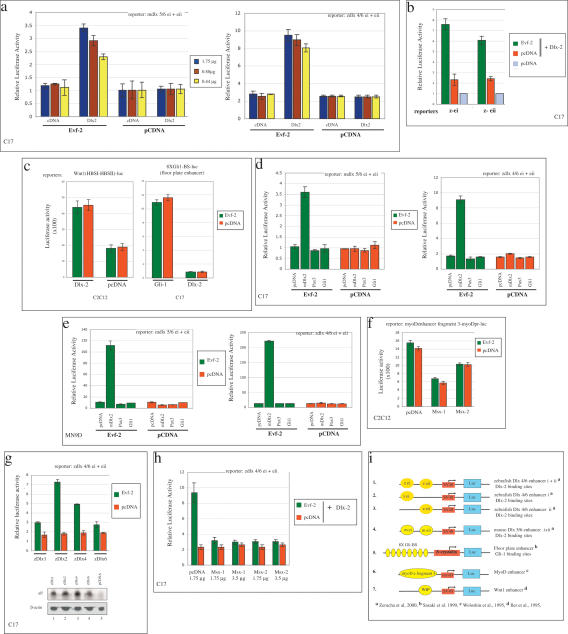
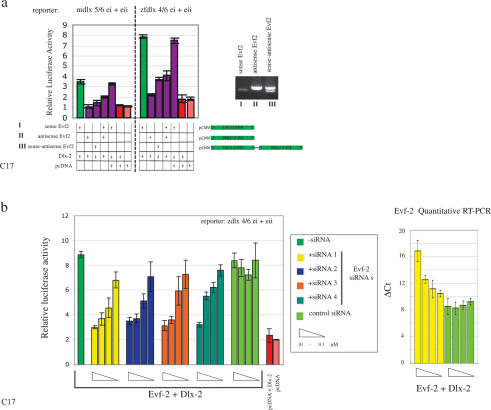
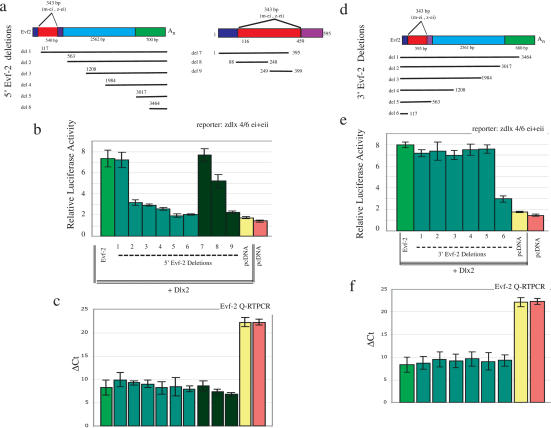
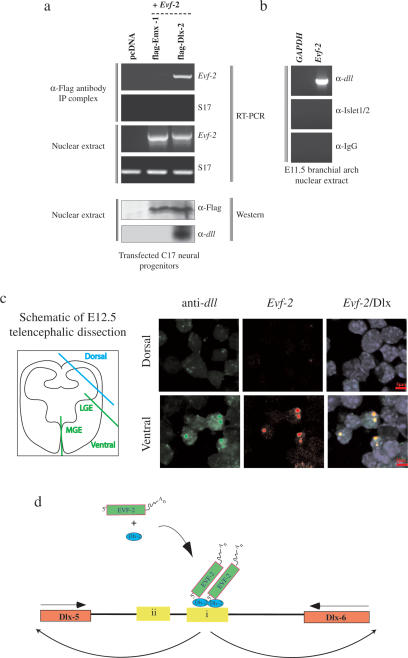
The identification of ultraconserved noncoding sequences in vertebrates has been associated with developmental regulators and DNA-binding proteins. One of the first of these was identified in the intergenic region between the Dlx-5 and Dlx-6 genes, members of the Dlx/dll homeodomain-containing protein family. In previous experiments, we showed that Sonic hedgehog treatment of forebrain neural explants results in the activation of Dlx-2 and the novel noncoding RNA (ncRNA), Evf-1. In this report, we show that the Dlx-5/6 ultraconserved region is transcribed to generate an alternatively spliced form of Evf-1, the ncRNA Evf-2. Evf-2 specifically cooperates with Dlx-2 to increase the transcriptional activity of the Dlx-5/6 enhancer in a target and homeodomain-specific manner. A stable complex containing the Evf-2 ncRNA and the Dlx-2 protein forms in vivo, suggesting that the Evf-2 ncRNA activates transcriptional activity by directly influencing Dlx-2 activity. These experiments identify a novel mechanism whereby transcription is controlled by the cooperative actions of an ncRNA and a homeodomain protein. The possibility that a subset of vertebrate ultraconserved regions may function at both the DNA and RNA level to control key developmental regulators may explain why ultraconserved sequences exhibit 90% or more conservation even after 450 million years of vertebrate evolution.
Figures
References
-
- Acampora D., Merlo G.R., Paleari L., Zerega B., Postiglione M.P., Mantero S., Bober E., Barbieri O., Simeone A., Levi G. Craniofacial, vestibular and bone defects in mice lacking the Distal-less related gene. Dlx5. Development. 1999;126:3795–3809. - PubMed
-
- Akhtar A. Dosage compensation: An intertwined world of RNA and chromatin remodeling. Curr. Opin. Genet. Dev. 2003;13:161–169. - PubMed
-
- Amrein H., Axel R. Genes expressed in neurons of adult male Drosophila. Cell. 1997;88:459–469. - PubMed
-
- Anderson S., Eisenstat D., Shi L., Rubenstein J. Interneuron migration from basal forebrain to neocortex: Dependence on Dlx genes. Science. 1997a;278:474–476. - PubMed
-
- Anderson S.A., Qiu M., Bulfone A., Eisenstat D.D., Meneses J., Pedersen R., Rubenstein J.L.R. Mutations of the homeobox genes Dlx-1 and Dlx-2 disrupt the striatal subventricular zone and differentiation of late born striatal neurons. Neuron. 1997b;19:27–37. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials