

Rapid phosphorylation of histone H2A.X following ionotropic glutamate receptor activation
- PMID: 16706843
- PMCID: PMC1534119
- DOI: 10.1111/j.1460-9568.2006.04768.x
Rapid phosphorylation of histone H2A.X following ionotropic glutamate receptor activation
Abstract
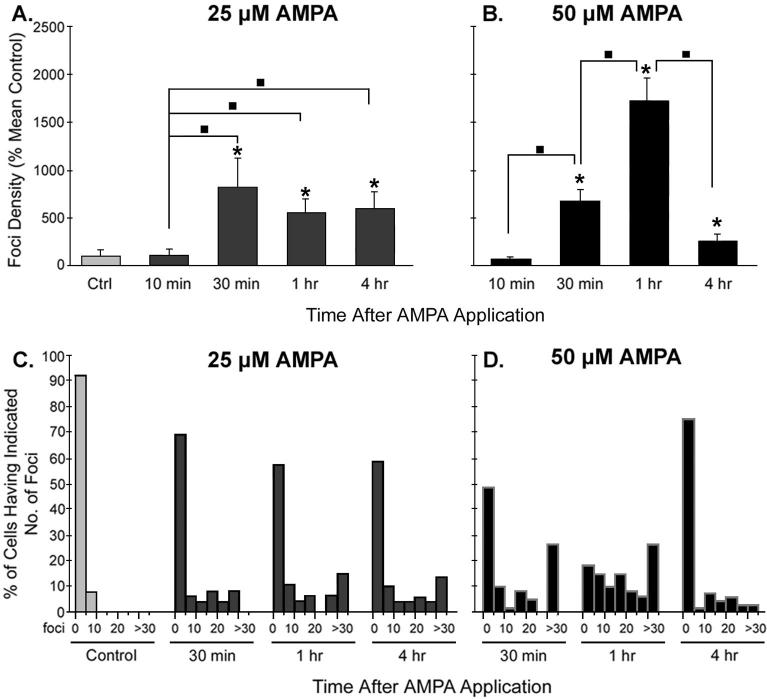
Excessive activation of ionotropic glutamate receptors increases oxidative stress, contributing to the neuronal death observed following neurological insults such as ischemia and seizures. Post-translational histone modifications may be key mediators in the detection and repair of damage resulting from oxidative stress, including DNA damage, and may thus affect neuronal survival in the aftermath of insults characterized by excessive glutamate release. In non-neuronal cells, phosphorylation of histone variant H2A.X (termed gamma-H2AX) occurs rapidly following DNA double-strand breaks. We investigated gamma-H2AX formation in rat cortical neurons (days in vitro 14) following activation of N-methyl-D-aspartate (NMDA) or alpha-amino-3-hydroxyl-5-methyl-4-isoxazolepropionic acid (AMPA)/kainate glutamate receptors using fluorescent immunohistochemical techniques. Moreover, we evaluated the co-localization of gamma-H2AX 'foci' with Mre11, a double-strand break repair protein, to provide further evidence for the activation of this DNA damage response pathway. Here we show that minimally cytotoxic stimulation of ionotropic glutamate receptors was sufficient to evoke gamma-H2AX in neurons, and that NMDA-induced gamma-H2AX foci formation was attenuated by pretreatment with the antioxidant, Vitamin E, and the intracellular calcium chelator, BAPTA-AM. Moreover, a subset of gamma-H2AX foci co-localized with Mre11, indicating that at least a portion of gamma-H2AX foci is damage dependent. The extent of gamma-H2AX induction following glutamate receptor activation corresponded to the increases we observed following conventional DNA damaging agents [i.e. non-lethal doses of gamma-radiation (1 Gy) and hydrogen peroxide (10 microm)]. These data suggest that insults not necessarily resulting in neuronal death induce the DNA damage-evoked chromatin modification, gamma-H2AX, and implicate a role for histone alterations in determining neuronal vulnerability following neurological insults.
Figures








References
-
- Balajee AS, Geard CR. Replication protein A and gamma-H2AX foci assembly is triggered by cellular response to DNA double-strand breaks. Exp. Cell Res. 2004;300:320–334. - PubMed
-
- Barnham KJ, Masters CL, Bush AI. Neurodegenerative diseases and oxidative stress. Nat. Rev. Drug Discov. 2004;3:205–214. - PubMed
-
- Bassing CH, Alt FW. H2AX may function as an anchor to hold broken chromosomal DNA ends in close proximity. Cell Cycle. 2004;3:149–153. - PubMed
-
- Blaisdell JO, Harrison L, Wallace SS. Base excision repair processing of radiation-induced clustered DNA lesions. Radiat. Prot. Dosimetry. 2001;97:25–31. - PubMed
-
- Brooks PJ. DNA repair in neural cells: basic science and clinical implications. Mutat. Res. 2002;509:93–108. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
