

Tests of rRNA hybridization to microarrays suggest that hybridization characteristics of oligonucleotide probes for species discrimination cannot be predicted
- PMID: 16707658
- PMCID: PMC1463897
- DOI: 10.1093/nar/gkl133
Tests of rRNA hybridization to microarrays suggest that hybridization characteristics of oligonucleotide probes for species discrimination cannot be predicted
Abstract
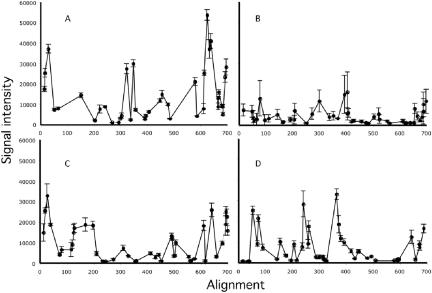
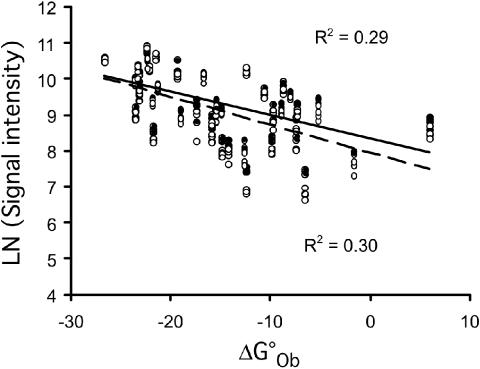
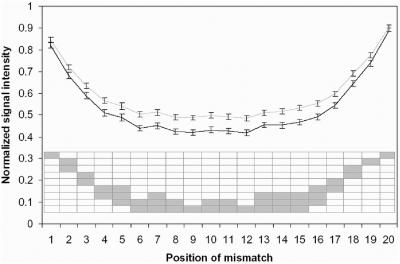
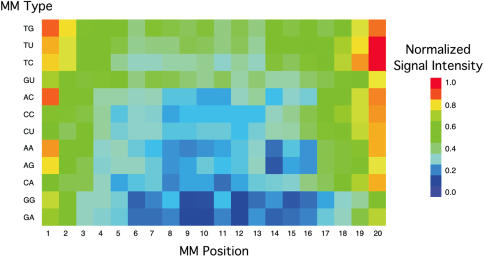
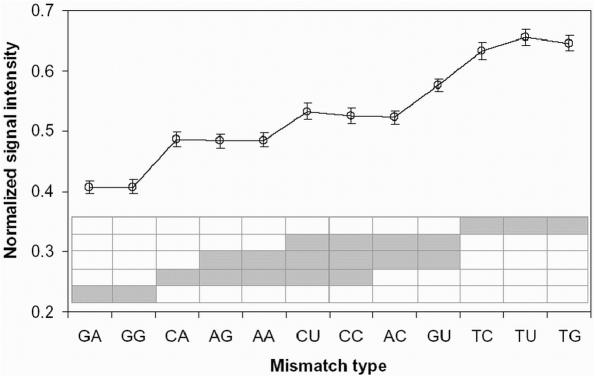
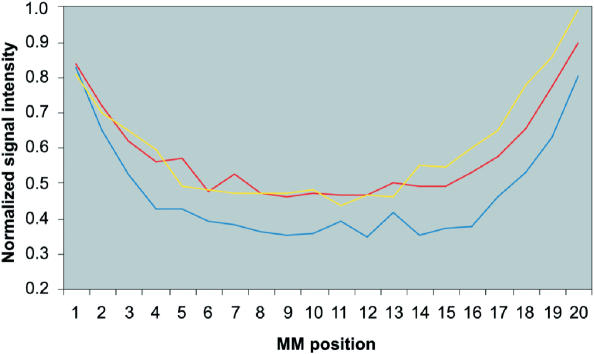
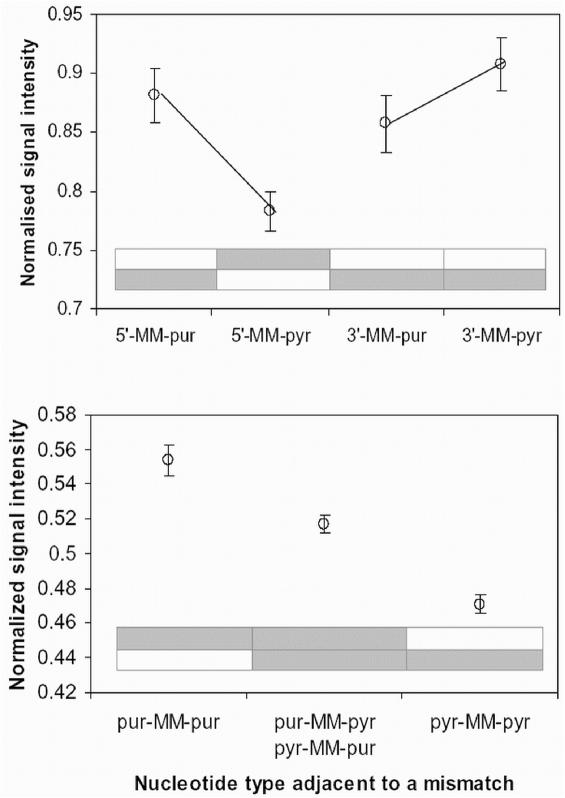
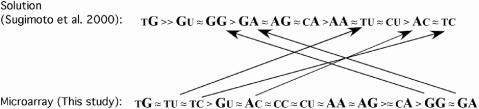
Hybridization of rRNAs to microarrays is a promising approach for prokaryotic and eukaryotic species identification. Typically, the amount of bound target is measured by fluorescent intensity and it is assumed that the signal intensity is directly related to the target concentration. Using thirteen different eukaryotic LSU rRNA target sequences and 7693 short perfect match oligonucleotide probes, we have assessed current approaches for predicting signal intensities by comparing Gibbs free energy (DeltaG degrees) calculations to experimental results. Our evaluation revealed a poor statistical relationship between predicted and actual intensities. Although signal intensities for a given target varied up to 70-fold, none of the predictors were able to fully explain this variation. Also, no combination of different free energy terms, as assessed by principal component and neural network analyses, provided a reliable predictor of hybridization efficiency. We also examined the effects of single-base pair mismatch (MM) (all possible types and positions) on signal intensities of duplexes. We found that the MM effects differ from those that were predicted from solution-based hybridizations. These results recommend against the application of probe design software tools that use thermodynamic parameters to assess probe quality for species identification. Our results imply that the thermodynamic properties of oligonucleotide hybridization are by far not yet understood.
Figures
References
-
- DeSantis T.Z., Stone C.E., Murray S.R., Moberg J.P., Andersen G.L. Rapid quantification and taxonomic classification of environmental DNA from both prokaryotic and eukaryotic origins using a microarray. FEMS Microbiol. Lett. 2005;245:271–278. - PubMed
-
- Wilson W.J., Strout C.L., DeSantis T.Z., Stilwell J.L., Carrano A.V., Andersen G.L. Sequence-specific identification of 18 pathogenic microorganisms using microarray technology. Mol. Cell. Probes. 2002;16:119–127. - PubMed
-
- Liu W.T., Mirzabekov A.D., Stahl D.A. Optimization of an oligonucleotide microchip for microbial identification studies: a non-equilibrium dissociation approach. Environ. Microbiol. 2001;3:619–629. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
