

Single-channel analysis of a point mutation of a conserved serine residue in the S2 ligand-binding domain of the NR2A NMDA receptor subunit
- PMID: 16709630
- PMCID: PMC1817770
- DOI: 10.1113/jphysiol.2006.112193
Single-channel analysis of a point mutation of a conserved serine residue in the S2 ligand-binding domain of the NR2A NMDA receptor subunit
Abstract
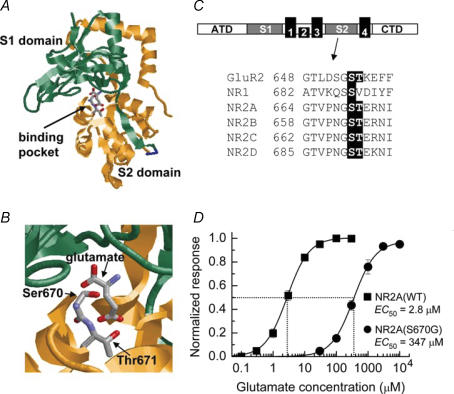
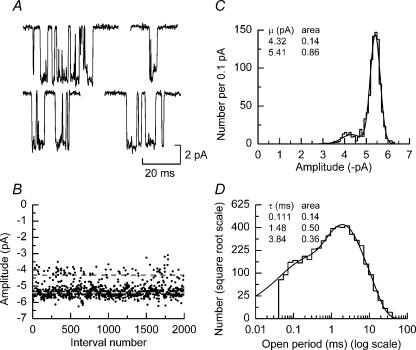
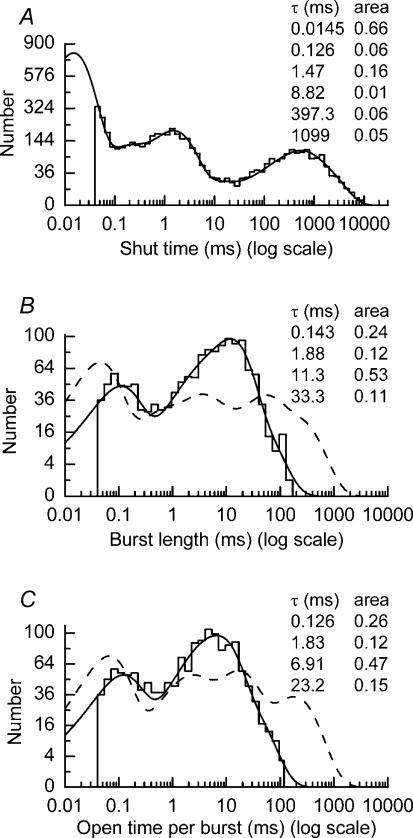
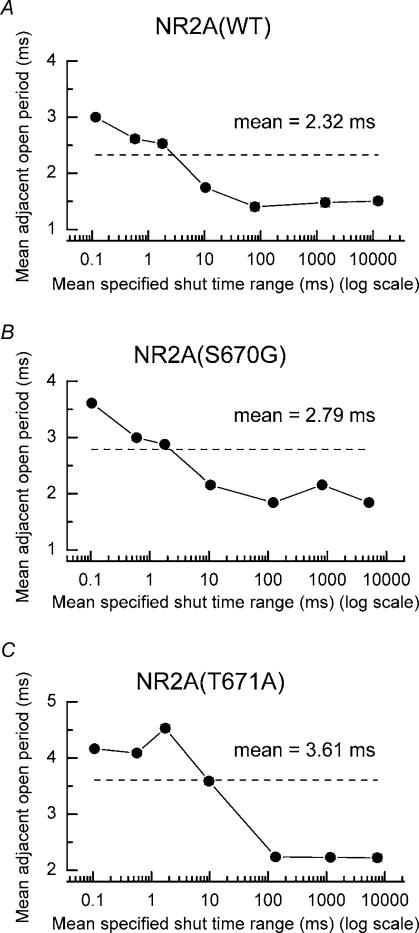
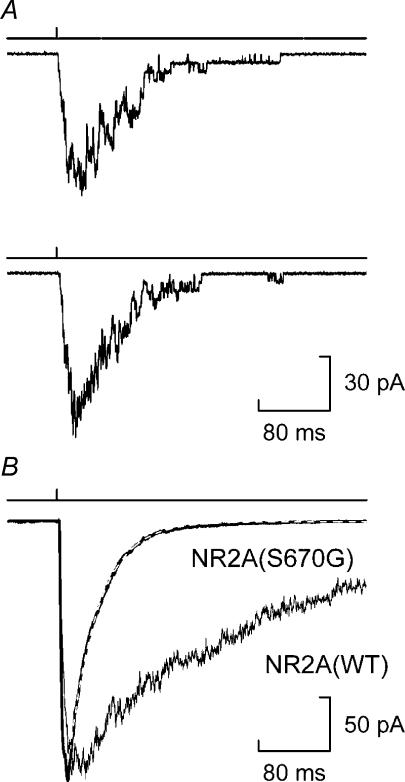
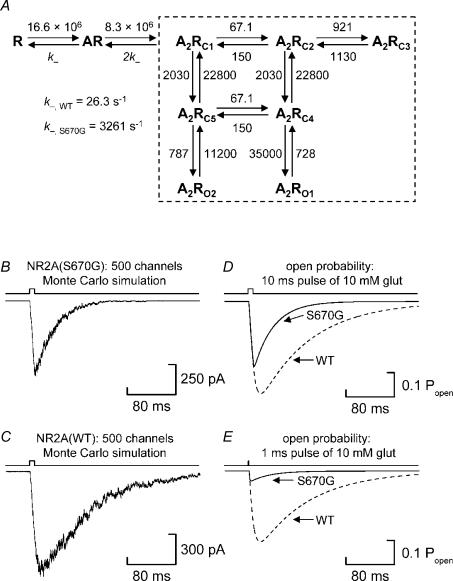
We have examined the function of a conserved serine residue (Ser670) in the S2 ligand-binding region of the NR2A N-methyl-d-aspartate (NMDA) receptor subunit, using recombinant NR1/NR2A receptors expressed in Xenopus laevis oocytes. Mutation of Ser670 to glycine (S670G) in NR2A reduced the potency of glutamate by 124-fold. Single-channel conductance and the duration of apparent open periods of NR2A(S670G) receptor mutants were, however, indistinguishable from wild-type NMDA receptors. NR1/NR2A(S670G) shut-time distributions were best described by a mixture of six exponential components, and the four shortest shut intervals of each distribution were considered to occur within a channel activation (burst). Bursts of single-channel openings were fitted with a mixture of four exponential components. The longest two components carried the majority of the charge transfer and had mean durations of 9.6 +/- 0.5 and 29.6 +/- 1.5 ms. The overall channel open probability during a burst was high (mean, 0.83 +/- 0.06). Consistent with a shortening of NMDA receptor-channel burst lengths was the observation of an increased deactivation rate of macroscopic currents evoked by brief applications of glutamate to outside-out membrane patches. Correlations between shut times and adjacent open times were observed in all data records. Noticeably, shorter than average openings tended to occur next to long closed periods, whereas longer than average openings tended to occur next to short closings. Our single-channel data, together with modelling using a kinetic scheme to describe channel activations, support our hypothesis that the S670G point mutation reduces the dwell time of glutamate in its binding site.
Figures

Comment in
-
Glutamate unbinding reveals new insights into NMDA receptor activation.J Physiol. 2006 Jul 15;574(Pt 2):329. doi: 10.1113/jphysiol.2006.114017. Epub 2006 May 25. J Physiol. 2006. PMID: 16728444 Free PMC article. No abstract available.
References
-
- Chen PE, Geballe MT, Stansfeld PJ, Johnston AR, Yuan H, Jacob AL, Snyder JP, Traynelis SF, Wyllie DJA. Structural features of the glutamate binding site in recombinant NR1/NR2A N-methyl-d-aspartate receptors determined by site-directed mutagenesis and molecular modeling. Mol Pharmacol. 2005;67:1470–1484. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources