

A unique strategy for mRNA cap methylation used by vesicular stomatitis virus
- PMID: 16709677
- PMCID: PMC1482520
- DOI: 10.1073/pnas.0509821103
A unique strategy for mRNA cap methylation used by vesicular stomatitis virus
Abstract
Nonsegmented negative-sense (nsNS) RNA viruses typically replicate within the host cell cytoplasm and do not have access to the host mRNA capping machinery. These viruses have evolved a unique mechanism for mRNA cap formation in that the guanylyltransferase transfers GDP rather than GMP onto the 5' end of the RNA. Working with vesicular stomatitis virus (VSV), a prototype nsNS RNA virus, we now provide genetic and biochemical evidence that its mRNA cap methylase activities are also unique. Using recombinant VSV, we determined the function in mRNA cap methylation of a predicted binding site in the polymerase for the methyl donor, S-adenosyl-l-methionine. We found that amino acid substitutions to this site disrupted methylation at the guanine-N-7 (G-N-7) position or at both the G-N-7 and ribose-2'-O (2'-O) positions of the mRNA cap. These studies provide genetic evidence that the two methylase activities share an S-adenosyl-l-methionine-binding site and show that, in contrast to other cap methylation reactions, methylation of the G-N-7 position is not required for 2'-O methylation. These findings suggest that VSV evolved an unusual strategy of mRNA cap methylation that may be shared by other nsNS RNA viruses.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
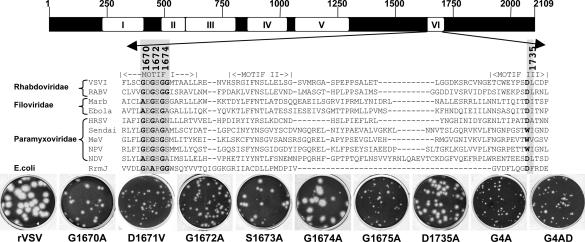
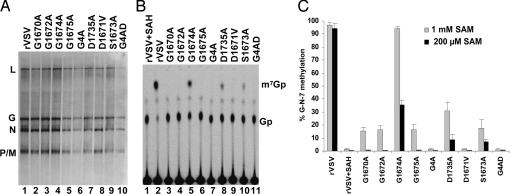
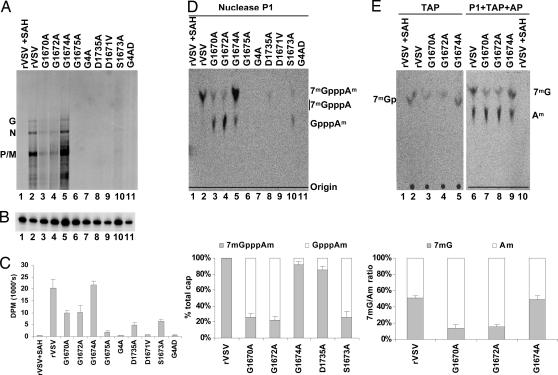
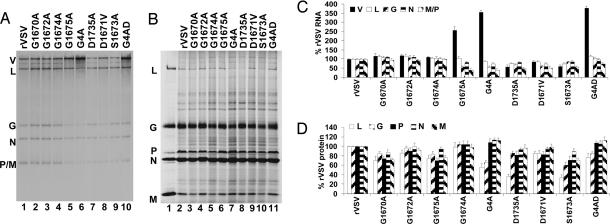
References
-
- Muthukrishnan S., Morgan M., Banerjee A. K., Shatkin A. J. Biochemistry. 1976;15:5761–5768. - PubMed
-
- Furuichi Y., LaFiandra A., Shatkin A. J. Nature. 1977;266:235–239. - PubMed
-
- Shuman S. Prog. Nucleic Acid Res. Mol. Biol. 2001;66:1–40. - PubMed
-
- Lima C. D., Wang L. K., Shuman S. Cell. 1999;99:533–543. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
