

Predicting RNA pseudoknot folding thermodynamics
- PMID: 16709732
- PMCID: PMC1463895
- DOI: 10.1093/nar/gkl346
Predicting RNA pseudoknot folding thermodynamics
Abstract
Based on the experimentally determined atomic coordinates for RNA helices and the self-avoiding walks of the P (phosphate) and C4 (carbon) atoms in the diamond lattice for the polynucleotide loop conformations, we derive a set of conformational entropy parameters for RNA pseudoknots. Based on the entropy parameters, we develop a folding thermodynamics model that enables us to compute the sequence-specific RNA pseudoknot folding free energy landscape and thermodynamics. The model is validated through extensive experimental tests both for the native structures and for the folding thermodynamics. The model predicts strong sequence-dependent helix-loop competitions in the pseudoknot stability and the resultant conformational switches between different hairpin and pseudoknot structures. For instance, for the pseudoknot domain of human telomerase RNA, a native-like and a misfolded hairpin intermediates are found to coexist on the (equilibrium) folding pathways, and the interplay between the stabilities of these intermediates causes the conformational switch that may underlie a human telomerase disease.
Figures



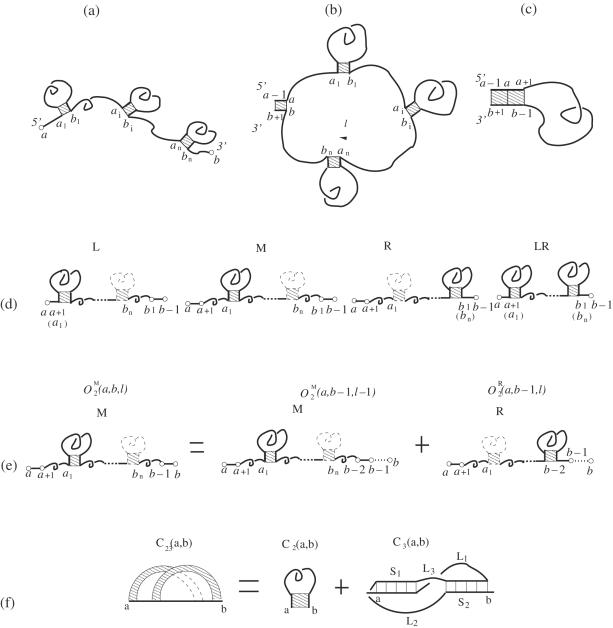

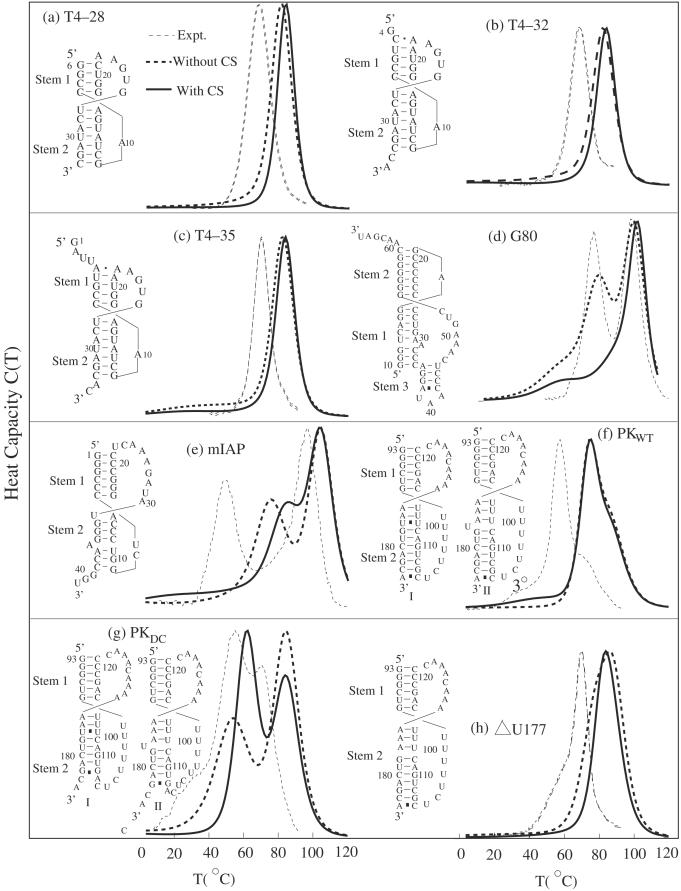

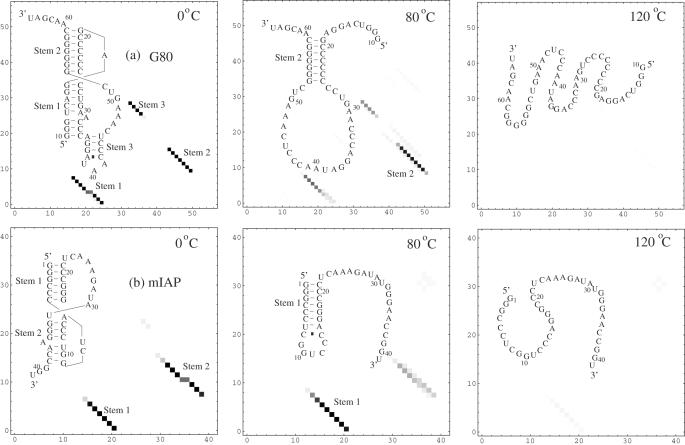


References
-
- Pleij C.W.A., Bosch L. RNA pseudoknots—structure, detection, and prediction. Meth. Enzymol. 1989;180:289–303. - PubMed
-
- Pleij C.W.A. Pseudoknots—a new motif in the RNA game. Trends Biochem. Sci. 1990;15:143–147. - PubMed
-
- Tinoco I., Jr, Puglisi J.D., Wyatt J.R. RNA Folding. In: Eckstein F., Lilley D., editors. Nucleic Acids and Molecular Biology. vol. 4. Berlin: Springer-Verlag press; 1990. pp. 205–226.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
