

Crystal structure of CHP2 complexed with NHE1-cytosolic region and an implication for pH regulation
- PMID: 16710297
- PMCID: PMC1478177
- DOI: 10.1038/sj.emboj.7601145
Crystal structure of CHP2 complexed with NHE1-cytosolic region and an implication for pH regulation
Abstract
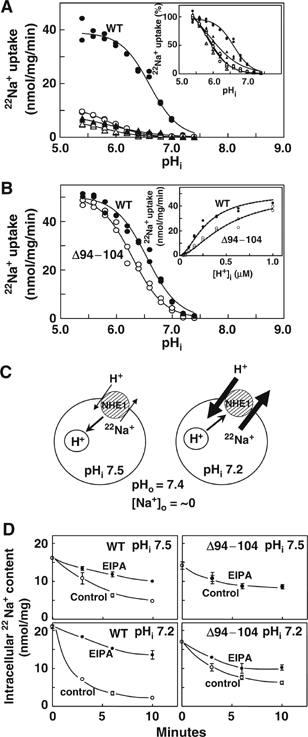
The plasma membrane Na+/H+ exchangers (NHE) require calcineurin B homologous protein (CHP) as an obligatory binding partner for ion transport. Here, we report the first crystal structure of CHP (CHP2 isoform) in complex with its binding domain in NHE1. We show that the cytoplasmic alpha-helix of NHE1 is inserted into the hydrophobic cleft formed by N- and C-lobes of CHP2 and that the size and shape of this crevice together with hydrogen bond formation at multiple positions assure a high degree of specificity for interaction with NHE members. Structure-based mutagenesis revealed the importance of hydrophobic interactions between CHP/NHE1 for the function of NHE1. Furthermore, the crystal structure shows the existence of a protruding CHP-unique region, and deletion of this region in CHP2 inhibited the NHE1 activity by inducing the acidic shift of intracellular pH dependence, while preserving interaction with NHE1. These findings suggest that CHP serves as an obligatory subunit that is required both for supporting the basic activity and regulating the pH-sensing of NHE1 via interactions between distinct parts of these proteins.
Figures





References
-
- Ames JB, Ishima R, Tanaka T, Gordon JI, Stryer L, Ikura M (1997) Molecular mechanics of calcium-myristoyl switches. Nature 389: 198–202 - PubMed
-
- An WF, Bowlby MR, Betty M, Cao J, Ling HP, Mendoza G, Hinson JW, Mattsson KI, Strassle BW, Trimmer JS, Rhodes KJ (2000) Modulation of A-type potassium channels by a family of calcium sensors. Nature 403: 553–556 - PubMed
-
- Aronson PS, Nee J, Suhm MA (1982) Modifier role of internal H+ in activating the Na+-H+ exchanger in renal microvillus membrane vesicles. Nature 299: 161–163 - PubMed
-
- Barroso MR, Bernd KK, DeWitt ND, Chang A, Mills K, Sztul ES (1996) A novel Ca2+-binding protein, p22, is required for constitutive membrane traffic. J Biol Chem 271: 10183–10187 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
