

Production and characterization of vaccines based on flaviviruses defective in replication
- PMID: 16712897
- PMCID: PMC2430078
- DOI: 10.1016/j.virol.2006.04.003
Production and characterization of vaccines based on flaviviruses defective in replication
Abstract
To develop new vaccine candidates for flavivirus infections, we have engineered two flaviviruses, yellow fever virus (YFV) and West Nile virus (WNV), that are deficient in replication. These defective pseudoinfectious viruses (PIVs) lack a functional copy of the capsid (C) gene in their genomes and are incapable of causing spreading infection upon infection of cells both in vivo and in vitro. However, they produce extracellular E protein in form of secreted subviral particles (SVPs) that are known to be an effective immunogen. PIVs can be efficiently propagated in trans-complementing cell lines making high levels of C or all three viral structural proteins. PIVs derived from YFV and WNV, demonstrated very high safety and immunization produced high levels of neutralizing antibodies and protective immune response. Such defective flaviviruses can be produced in large scale under low biocontainment conditions and should be useful for diagnostic or vaccine applications.
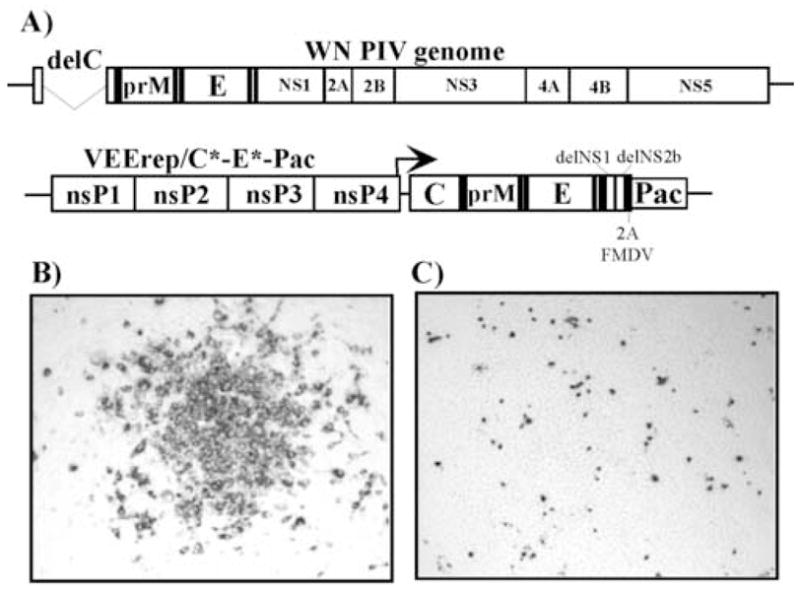
Figures
Similar articles
-
Efficient, trans-complementing packaging systems for chimeric, pseudoinfectious dengue 2/yellow fever viruses.Virology. 2010 Apr 25;400(1):8-17. doi: 10.1016/j.virol.2009.12.015. Virology. 2010. PMID: 20137799 Free PMC article.
-
Production of pseudoinfectious yellow fever virus with a two-component genome.J Virol. 2007 Nov;81(21):11737-48. doi: 10.1128/JVI.01112-07. Epub 2007 Aug 22. J Virol. 2007. PMID: 17715227 Free PMC article.
-
Replication-Defective West Nile Virus with NS1 Deletion as a New Vaccine Platform for Flavivirus.J Virol. 2019 Aug 13;93(17):e00720-19. doi: 10.1128/JVI.00720-19. Print 2019 Sep 1. J Virol. 2019. PMID: 31189715 Free PMC article.
-
Preclinical and clinical development of YFV 17D-based chimeric vaccines against dengue, West Nile and Japanese encephalitis viruses.Vaccine. 2010 Jan 8;28(3):632-49. doi: 10.1016/j.vaccine.2009.09.098. Epub 2009 Oct 4. Vaccine. 2010. PMID: 19808029 Review.
-
Yellow fever vector live-virus vaccines: West Nile virus vaccine development.Trends Mol Med. 2001 Aug;7(8):350-4. doi: 10.1016/s1471-4914(01)02048-2. Trends Mol Med. 2001. PMID: 11516995 Free PMC article. Review.
Cited by
-
Engineering the Japanese encephalitis virus RNA genome for the expression of foreign genes of various sizes: implications for packaging capacity and RNA replication efficiency.J Neurovirol. 2007 Dec;13(6):522-35. doi: 10.1080/13550280701684651. J Neurovirol. 2007. PMID: 18097884
-
West Nile virus genome with glycosylated envelope protein and deletion of alpha helices 1, 2, and 4 in the capsid protein is noninfectious and efficiently secretes subviral particles.J Virol. 2013 Dec;87(23):13063-9. doi: 10.1128/JVI.01552-13. Epub 2013 Sep 18. J Virol. 2013. PMID: 24049184 Free PMC article.
-
An Overview of Current Approaches Toward the Treatment and Prevention of West Nile Virus Infection.Methods Mol Biol. 2016;1435:249-91. doi: 10.1007/978-1-4939-3670-0_19. Methods Mol Biol. 2016. PMID: 27188563 Free PMC article.
-
Early production of type I interferon during West Nile virus infection: role for lymphoid tissues in IRF3-independent interferon production.J Virol. 2007 Sep;81(17):9100-8. doi: 10.1128/JVI.00316-07. Epub 2007 Jun 13. J Virol. 2007. PMID: 17567689 Free PMC article.
-
DENV2 Pseudoviral Particles with Unprocessed Capsid Protein Are Assembled and Infectious.Viruses. 2019 Dec 25;12(1):27. doi: 10.3390/v12010027. Viruses. 2019. PMID: 31881703 Free PMC article.
References
-
- Aberle JH, Aberle SW, Allison SL, Stiasny K, Ecker M, Mandl CW, Berger R, Heinz FX. A DNA immunization model study with constructs expressing the tick-borne encephalitis virus envelope protein E in different physical forms. J Immunol. 1999;163:6756–61. - PubMed
-
- Bredenbeek PJ, Kooi EA, Lindenbach B, Huijkman N, Rice CM, Spaan WJ. A stable full-length yellow fever virus cDNA clone and the role of conserved RNA elements in flavivirus replication. J Gen Virol. 2003;84:1261–8. - PubMed
-
- Colombage G, Hall R, Pavy M, Lobigs M. DNA-based and alphavirus-vectored immunisation with prM and E proteins elicits long-lived and protective immunity against the flavivirus, Murray Valley encephalitis virus. Virology. 1998;250:151–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
