

A bifunctional tRNA import receptor from Leishmania mitochondria
- PMID: 16714384
- PMCID: PMC1482498
- DOI: 10.1073/pnas.0510869103
A bifunctional tRNA import receptor from Leishmania mitochondria
Retraction in
-
Retraction for Goswami et al., A bifunctional tRNA import receptor from Leishmania mitochondria.Proc Natl Acad Sci U S A. 2025 Jul 29;122(30):e2517057122. doi: 10.1073/pnas.2517057122. Epub 2025 Jul 25. Proc Natl Acad Sci U S A. 2025. PMID: 40711922 Free PMC article. No abstract available.
Expression of concern in
-
Editorial Expression of Concern: A bifunctional tRNA import receptor from Leishmania mitochondria.Proc Natl Acad Sci U S A. 2010 May 18;107(20):9476. doi: 10.1073/pnas.1005260107. Proc Natl Acad Sci U S A. 2010. PMID: 28277630 Free PMC article. No abstract available.
Abstract
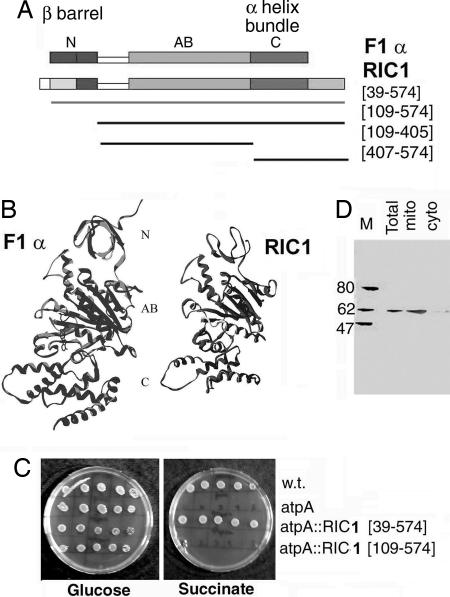
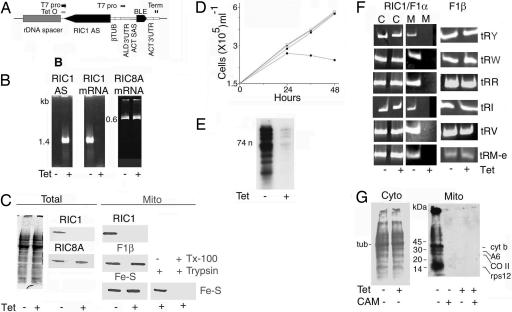
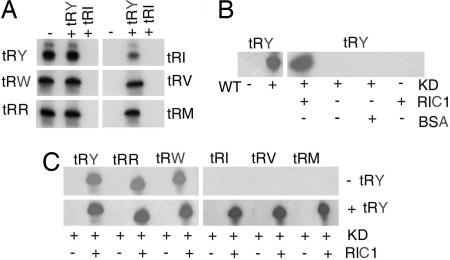
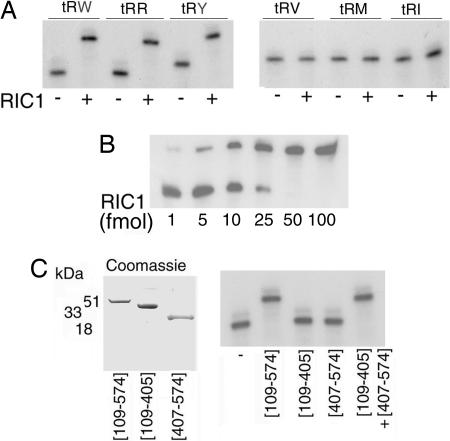
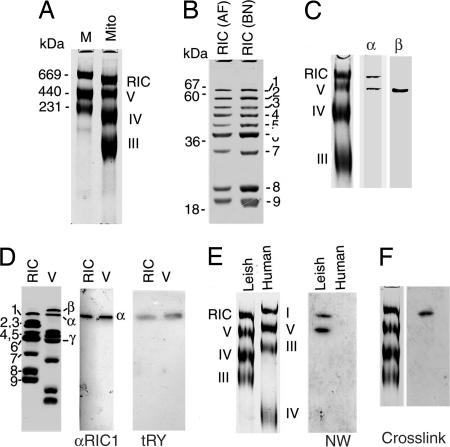
In kinetoplastid protozoa, import of cytosolic tRNAs into mitochondria occurs through tRNAs interacting with membrane-bound proteins, the identities of which are unknown. The inner membrane RNA import complex of Leishmania tropica contains multiple proteins and is active for import in vitro. RIC1, the largest subunit of this complex, is structurally homologous to the conserved alpha subunit of F1 ATP synthase. The RIC1 gene complemented an atpA mutation in Escherichia coli. Antisense-mediated knockdown of RIC1/F1alpha in Leishmania resulted in depletion of several mitochondrial tRNAs belonging to distinct subsets (types I and II) that interact cooperatively or antagonistically within the import complex. The knockdown-induced defect in import of type I tRNAs was rectified in a reconstituted system by purified RIC1/F1alpha alone, but recovery of type II tRNA import additionally required a type I tRNA. RIC1/F1alpha formed stable complexes with type I, but not type II, tRNAs through the cooperation of its nucleotide binding and C-terminal domains. Thus, RIC1/F1alpha is a type I tRNA import receptor. As expected of a bifunctional protein, RIC1/F1alpha is shared by both the import complex and by respiratory complex V. Alternative use of ancient respiratory proteins may have been an important step in the evolution of tRNA import.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
Similar articles
-
A mosaic of RNA binding and protein interaction motifs in a bifunctional mitochondrial tRNA import factor from Leishmania tropica.Nucleic Acids Res. 2008 Oct;36(17):5552-61. doi: 10.1093/nar/gkn536. Epub 2008 Aug 28. Nucleic Acids Res. 2008. PMID: 18755708 Free PMC article.
-
An RNA-binding respiratory component mediates import of type II tRNAs into Leishmania mitochondria.J Biol Chem. 2006 Sep 1;281(35):25270-7. doi: 10.1074/jbc.M604126200. Epub 2006 Jul 6. J Biol Chem. 2006. PMID: 16825195
-
The alpha-subunit of Leishmania F1 ATP synthase hydrolyzes ATP in presence of tRNA.J Biol Chem. 2006 Jul 14;281(28):18914-7. doi: 10.1074/jbc.C600089200. Epub 2006 May 30. J Biol Chem. 2006. PMID: 16735512
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Management of urinary stones by experts in stone disease (ESD 2025).Arch Ital Urol Androl. 2025 Jun 30;97(2):14085. doi: 10.4081/aiua.2025.14085. Epub 2025 Jun 30. Arch Ital Urol Androl. 2025. PMID: 40583613 Review.
Cited by
-
Mitochondrion-to-nucleus communication mediated by RNA export: a survey of potential mechanisms and players across eukaryotes.Biol Lett. 2024 Jul;20(7):20240147. doi: 10.1098/rsbl.2024.0147. Epub 2024 Jul 10. Biol Lett. 2024. PMID: 38982851 Free PMC article. Review.
-
Necessary and sufficient factors for the import of transfer RNA into the kinetoplast mitochondrion.EMBO Rep. 2007 Jun;8(6):589-95. doi: 10.1038/sj.embor.7400979. Epub 2007 May 18. EMBO Rep. 2007. PMID: 17510656 Free PMC article.
-
Mitochondrial Genome Engineering: The Revolution May Not Be CRISPR-Ized.Trends Genet. 2018 Feb;34(2):101-110. doi: 10.1016/j.tig.2017.11.001. Epub 2017 Nov 24. Trends Genet. 2018. PMID: 29179920 Free PMC article. Review.
-
RNA editing in Trypanosoma brucei requires three different editosomes.Mol Cell Biol. 2008 Jan;28(1):122-30. doi: 10.1128/MCB.01374-07. Epub 2007 Oct 22. Mol Cell Biol. 2008. PMID: 17954557 Free PMC article.
-
The ins and outs of tRNA transport.EMBO Rep. 2007 Jun;8(6):547-9. doi: 10.1038/sj.embor.7400989. EMBO Rep. 2007. PMID: 17545994 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
