

Human telomeric sequence forms a hybrid-type intramolecular G-quadruplex structure with mixed parallel/antiparallel strands in potassium solution
- PMID: 16714449
- PMCID: PMC1464114
- DOI: 10.1093/nar/gkl348
Human telomeric sequence forms a hybrid-type intramolecular G-quadruplex structure with mixed parallel/antiparallel strands in potassium solution
Abstract
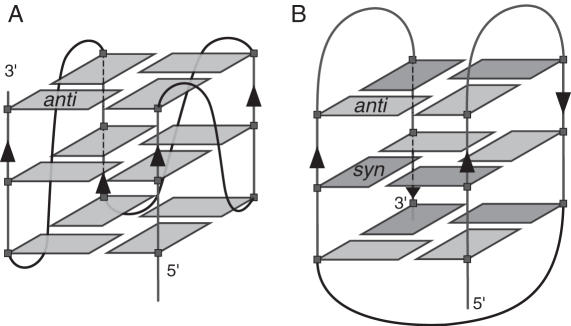
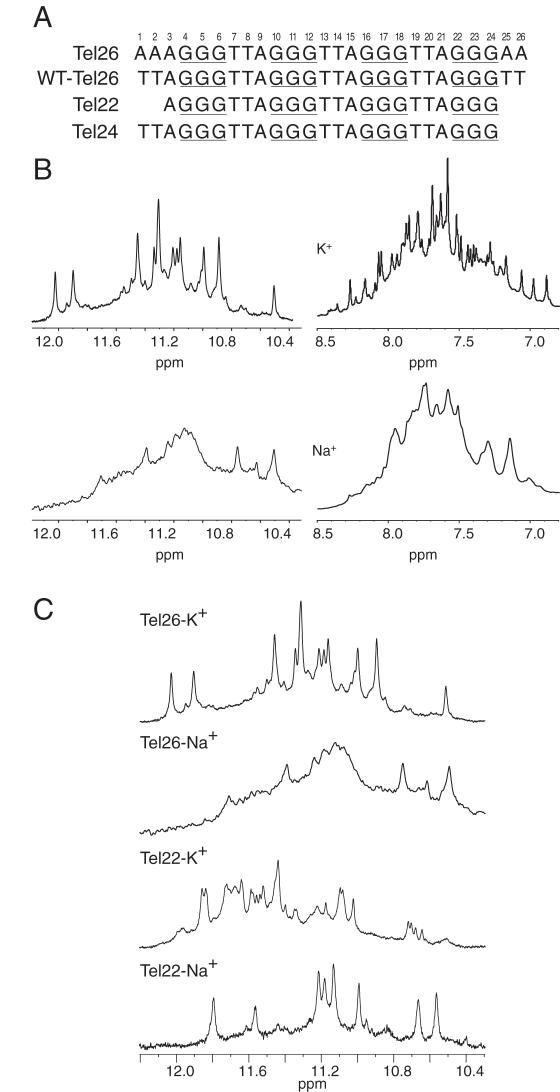
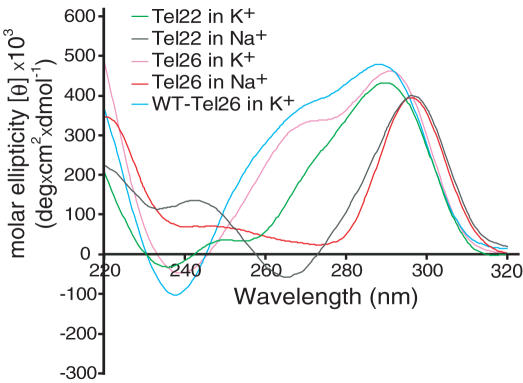
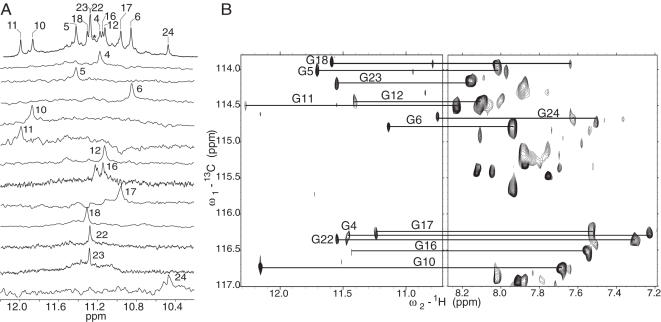
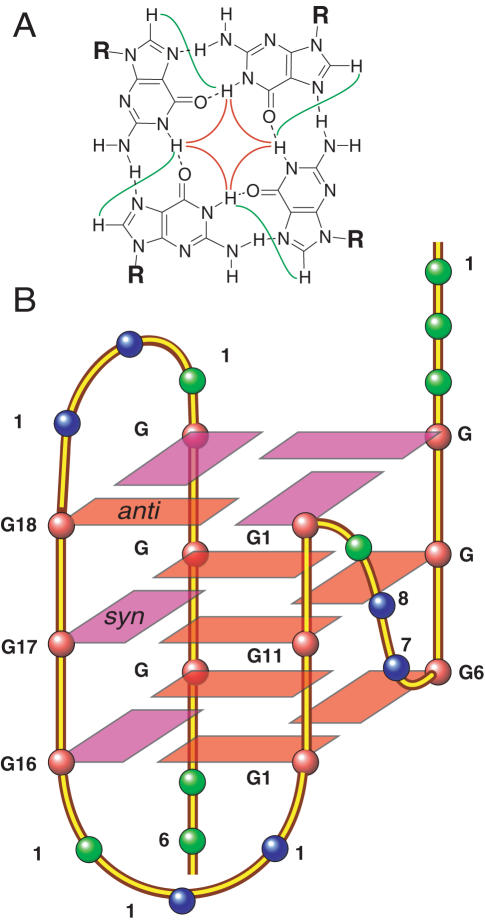
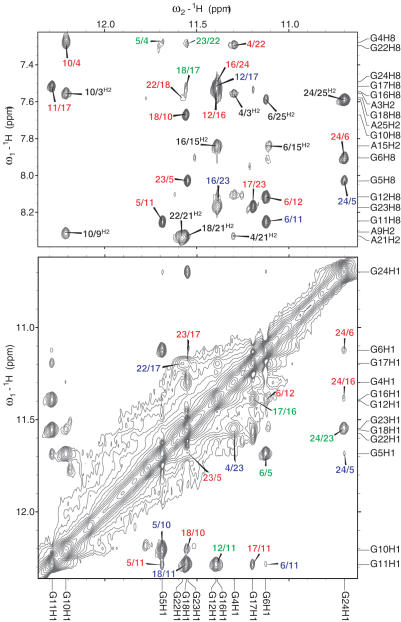
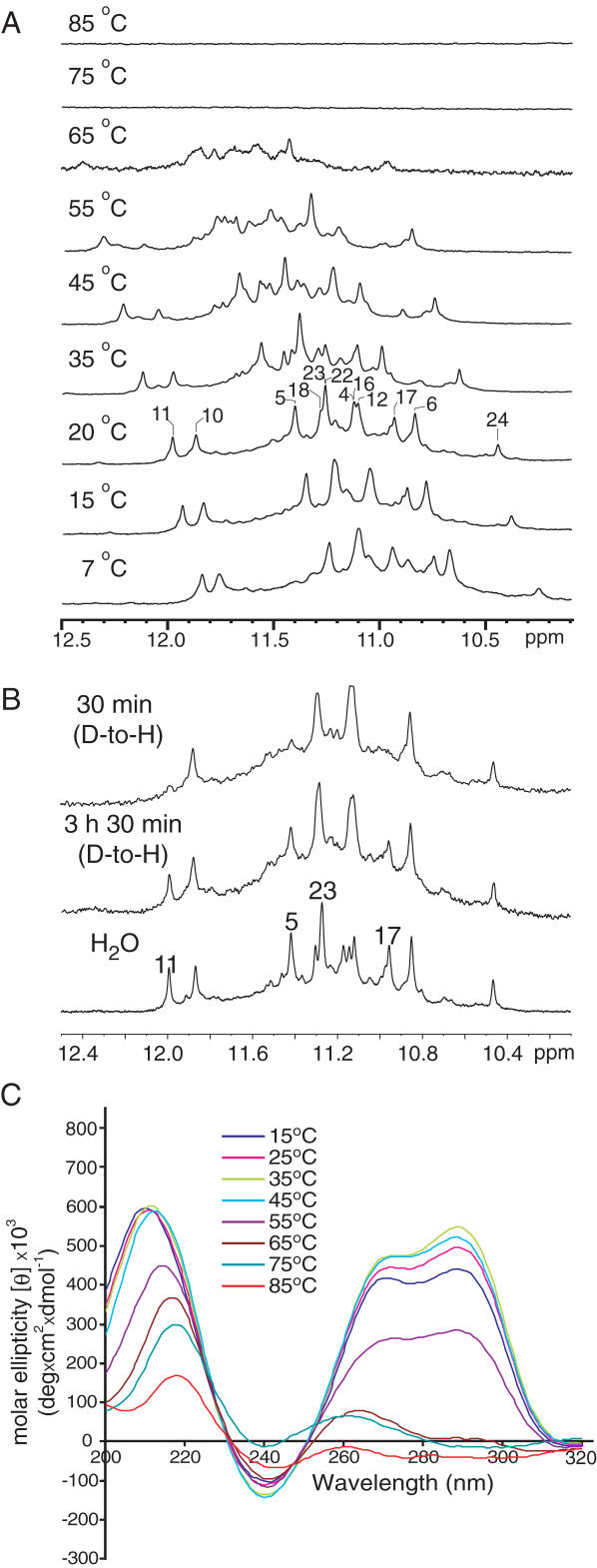
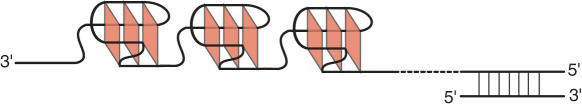
Human telomeric DNA consists of tandem repeats of the sequence d(TTAGGG). The formation and stabilization of DNA G-quadruplexes in the human telomeric sequence have been shown to inhibit the activity of telomerase, thus the telomeric DNA G-quadruplex has been considered as an attractive target for cancer therapeutic intervention. However, knowledge of the intact human telomeric G-quadruplex structure(s) formed under physiological conditions is a prerequisite for structure-based rational drug design. Here we report the folding structure of the human telomeric sequence in K+ solution determined by NMR. Our results demonstrate a novel, unprecedented intramolecular G-quadruplex folding topology with hybrid-type mixed parallel/antiparallel G-strands. This telomeric G-quadruplex structure contains three G-tetrads with mixed G-arrangements, which are connected consecutively with a double-chain-reversal side loop and two lateral loops, each consisting of three nucleotides TTA. This intramolecular hybrid-type telomeric G-quadruplex structure formed in K+ solution is distinct from those reported on the 22 nt Tel22 in Na+ solution and in crystalline state in the presence of K+, and appears to be the predominant conformation for the extended 26 nt telomeric sequence Tel26 in the presence of K+, regardless of the presence or absence of Na+. Furthermore, the addition of K+ readily converts the Na+-form conformation to the K+-form hybrid-type G-quadruplex. Our results explain all the reported experimental data on the human telomeric G-quadruplexes formed in the presence of K+, and provide important insights for understanding the polymorphism and interconversion of various G-quadruplex structures formed within the human telomeric sequence, as well as the effects of sequence and cations. This hybrid-type G-quadruplex topology suggests a straightforward pathway for the secondary structure formation with effective packing within the extended human telomeric DNA. The hybrid-type telomeric G-quadruplex is most likely to be of pharmacological relevance, and the distinct folding topology of this G-quadruplex suggests that it can be specifically targeted by G-quadruplex interactive small molecule drugs.
Figures



References
-
- Blackburn E.H. Telomere states and cell fates. Nature. 2000;408:53–56. - PubMed
-
- van Steensel B., Smogorzewska A., de Lange T. TRF2 protects human telomeres from end-to-end fusions. Cell. 1998;92:401–413. - PubMed
-
- Hackett J.A., Feldser D.M., Greider C.W. Telomere dysfunction increases mutation rate and genomic instability. Cell. 2001;106:275–286. - PubMed
-
- Makarov V.L., Hirose Y., Langmore J.P. Long G tails at both ends of human chromosomes suggest a C strand degradation mechanism for telomere shortening. Cell. 1997;88:657–666. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
