

Ultrafast dynamics in DNA: "fraying" at the end of the helix
- PMID: 16719468
- PMCID: PMC2528932
- DOI: 10.1021/ja0582105
Ultrafast dynamics in DNA: "fraying" at the end of the helix
Abstract
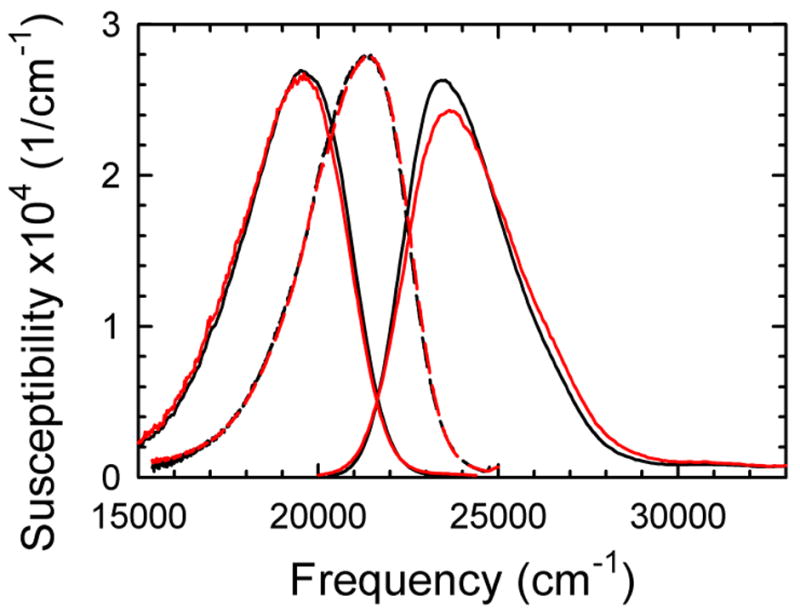
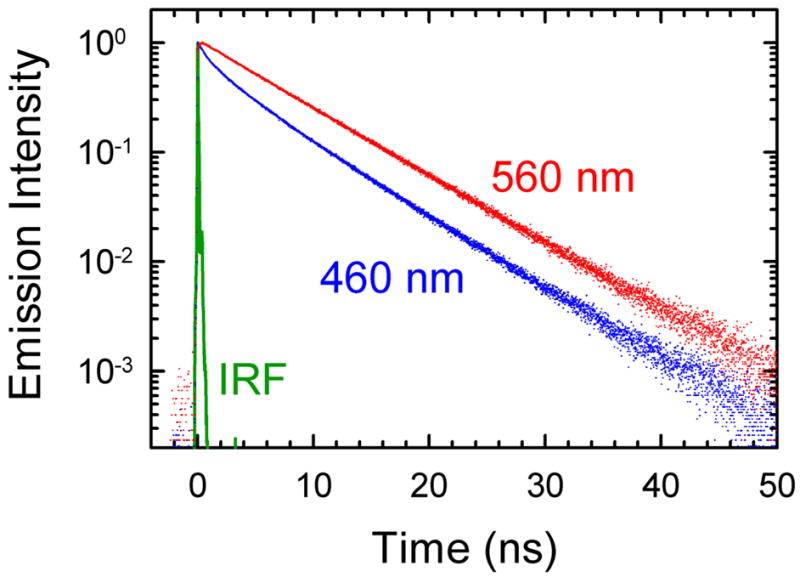
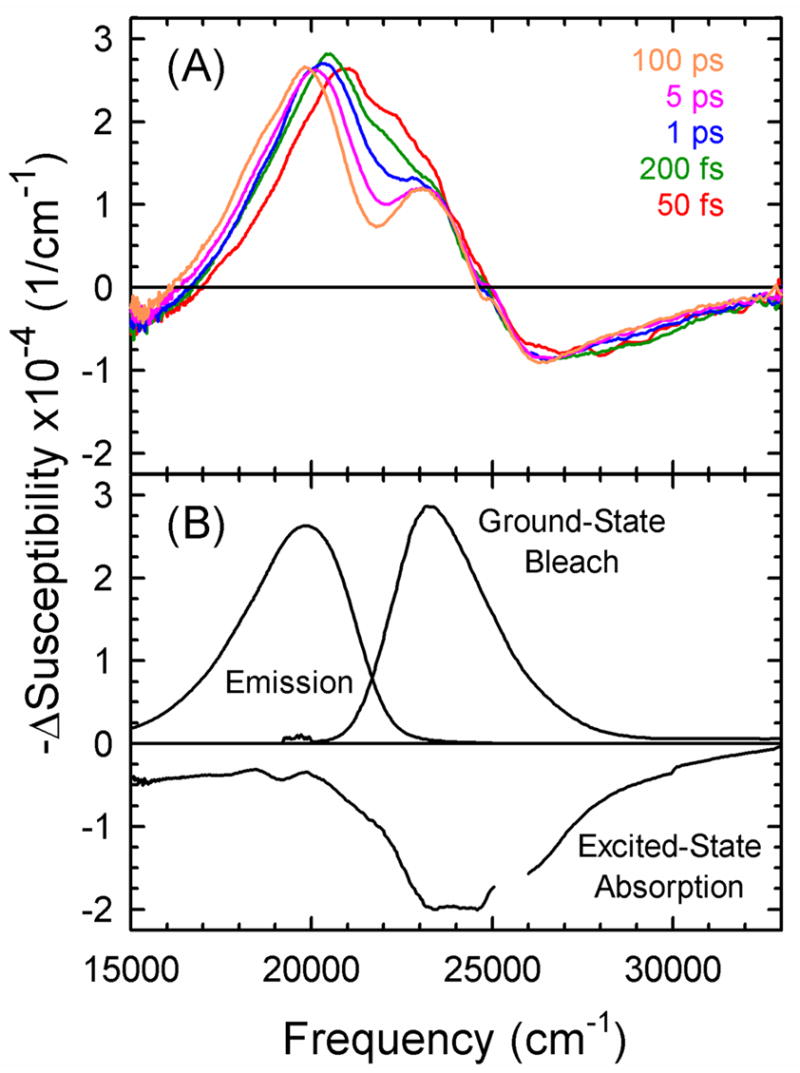
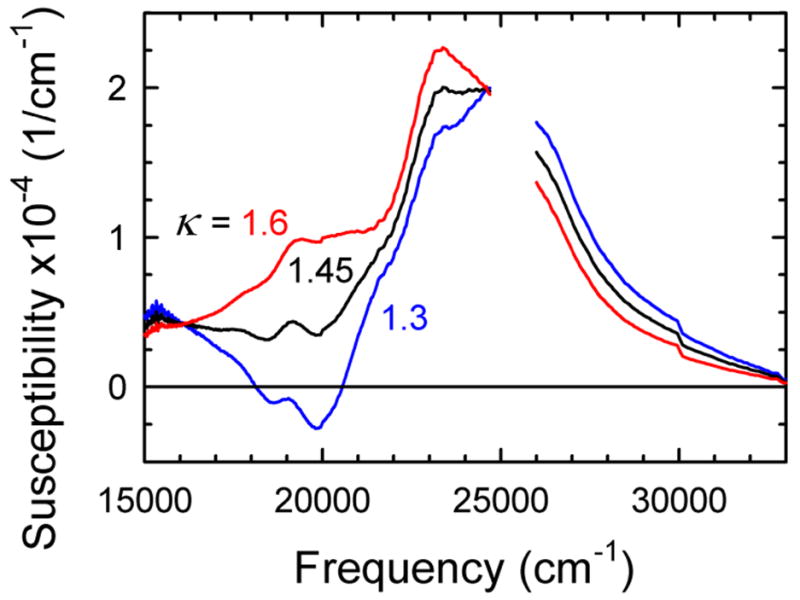
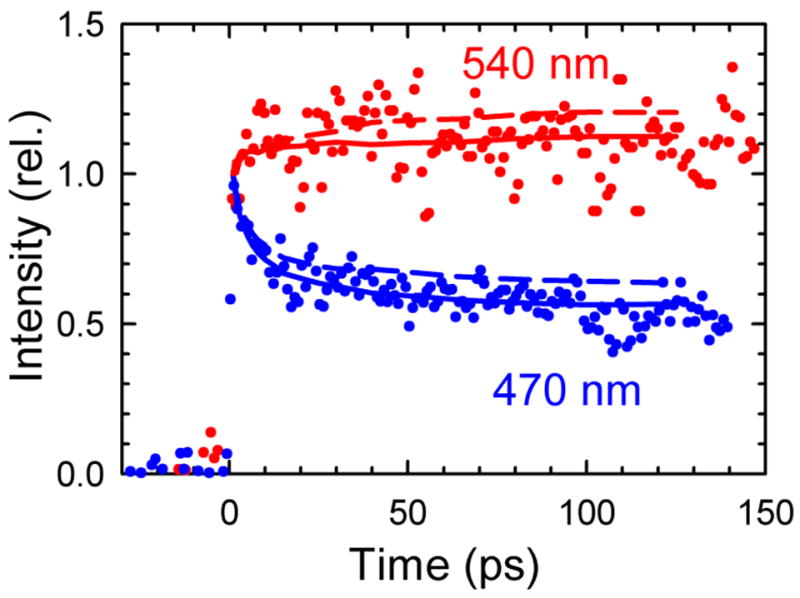
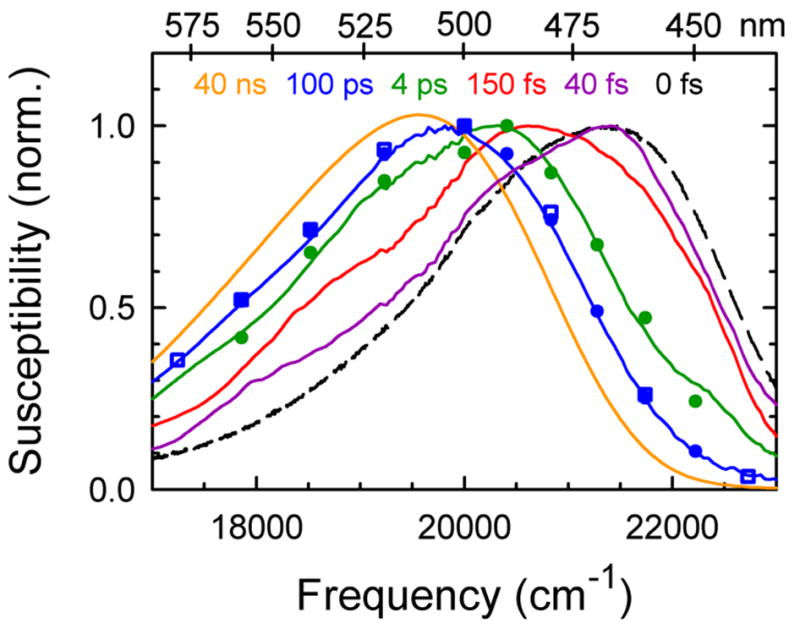
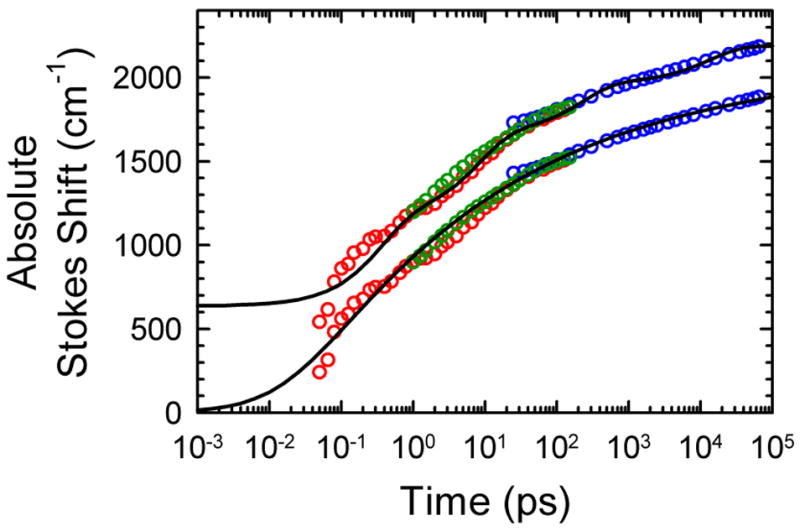
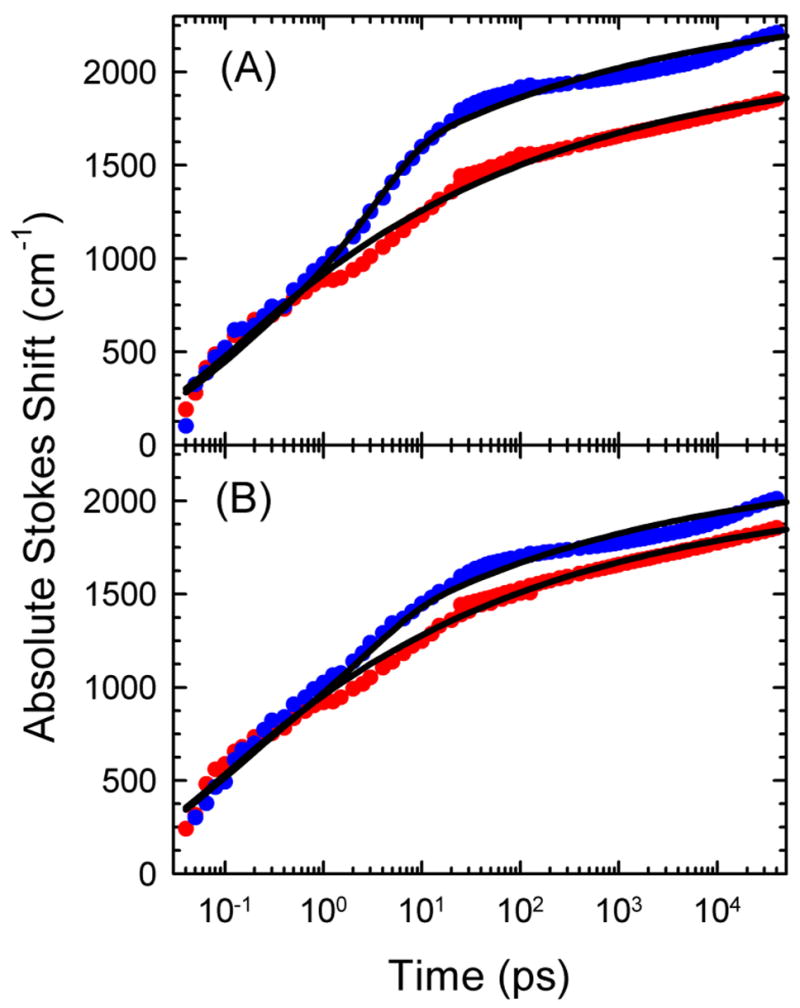
The dynamics of the electric fields in the interior of DNA are measured by using oligonucleotides in which a native base pair is replaced by a dye molecule (coumarin 102) whose emission spectrum is sensitive to the local electric field. Time-resolved measurements of the emission spectrum have been extended to a six decade time range (40 fs to 40 ns) by combining results from time-correlated photon counting, fluorescence up-conversion, and transient absorption. Recent results showed that when the reporter is placed in the center of the oligonucleotide, the dynamics are very broadly distributed over this entire time range and do not show specific time constants associated with individual processes (Andreatta, D.; et al. J. Am. Chem. Soc. 2005, 127, 7270). This paper examines an oligonucleotide with the reporter near its end. The broadly distributed relaxation seen before remains with little attenuation. In addition, a new relaxation with a well-defined relaxation time of 5 ps appears. This process is assigned to the rapid component of "fraying" at the end of the helix.
Figures
References
-
- Leroy JL, Kochoyan M, Huynhdinh T, Guéron M. J Mol Biol. 1988;200:223. - PubMed
-
- Nonin S, Leroy JL, Guéron M. Biochemistry. 1995;34:10652. - PubMed
-
- Holbrook SR, Kim SH. J Mol Biol. 1984;173:361. - PubMed
-
- Fujimoto BS, Willie ST, Reid BR, Schurr JM. J Mag Res B. 1995;106:64. - PubMed
-
- Borer PN, Laplante SR, Kumar A, Zanatta N, Martin A, Hakkinen A, Levy GC. Biochemistry. 1994;33:2441. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
