

Partition function and base pairing probabilities of RNA heterodimers
- PMID: 16722605
- PMCID: PMC1459172
- DOI: 10.1186/1748-7188-1-3
Partition function and base pairing probabilities of RNA heterodimers
Abstract
Background: RNA has been recognized as a key player in cellular regulation in recent years. In many cases, non-coding RNAs exert their function by binding to other nucleic acids, as in the case of microRNAs and snoRNAs. The specificity of these interactions derives from the stability of inter-molecular base pairing. The accurate computational treatment of RNA-RNA binding therefore lies at the heart of target prediction algorithms.
Methods: The standard dynamic programming algorithms for computing secondary structures of linear single-stranded RNA molecules are extended to the co-folding of two interacting RNAs.
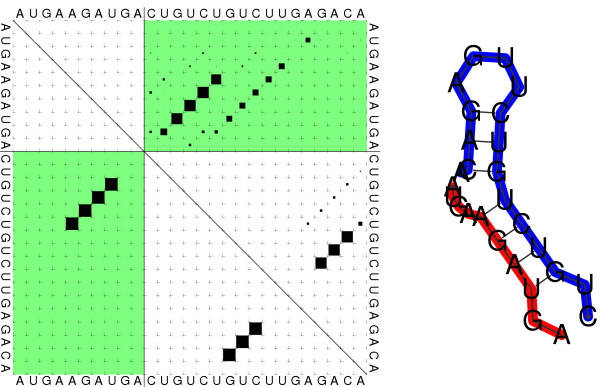
Results: We present a program, RNAcofold, that computes the hybridization energy and base pairing pattern of a pair of interacting RNA molecules. In contrast to earlier approaches, complex internal structures in both RNAs are fully taken into account. RNAcofold supports the calculation of the minimum energy structure and of a complete set of suboptimal structures in an energy band above the ground state. Furthermore, it provides an extension of McCaskill's partition function algorithm to compute base pairing probabilities, realistic interaction energies, and equilibrium concentrations of duplex structures.
Figures



References
LinkOut - more resources
Full Text Sources
Other Literature Sources
