

Mrc1 and Srs2 are major actors in the regulation of spontaneous crossover
- PMID: 16724109
- PMCID: PMC1500851
- DOI: 10.1038/sj.emboj.7601158
Mrc1 and Srs2 are major actors in the regulation of spontaneous crossover
Abstract
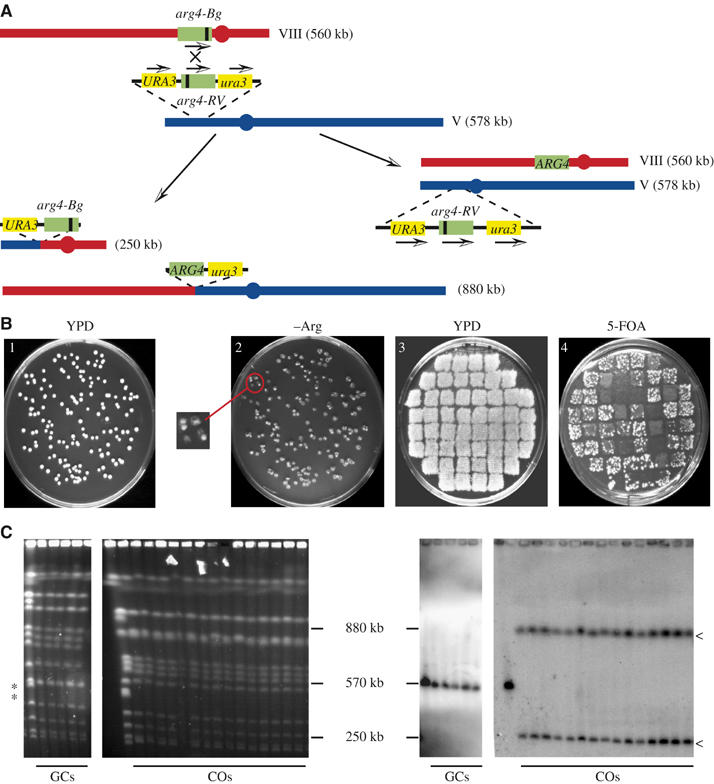
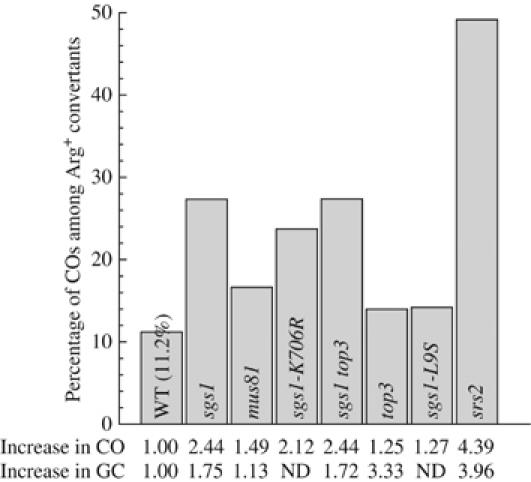
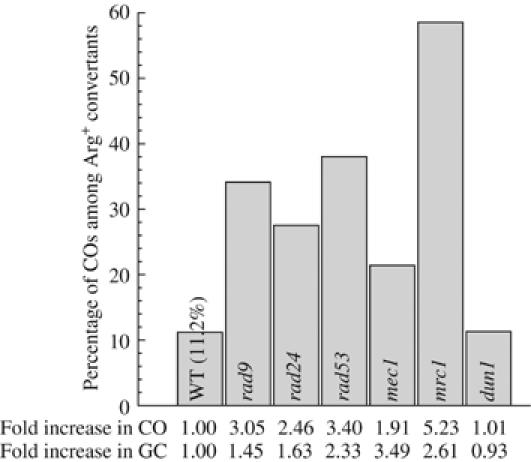
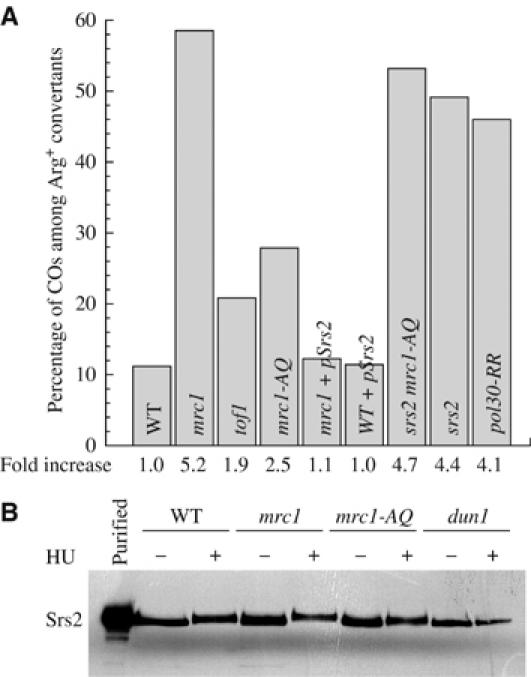
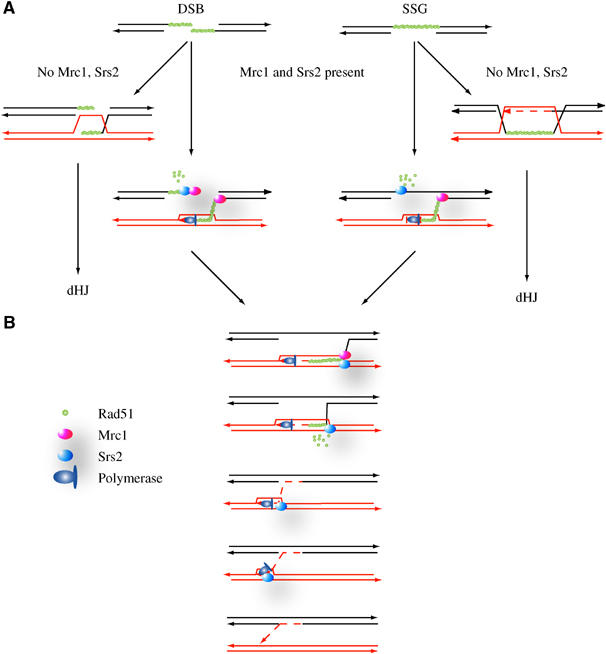
In vegetative cells, most recombination intermediates are metabolized without an association with a crossover (CO). The avoidance of COs allows for repair and prevents genomic rearrangements, potentially deleterious if the sequences involved are at ectopic locations. We have designed a system that permits to screen spontaneous intragenic recombination events in Saccharomyces cerevisiae and to investigate the CO outcome in different genetic contexts. We have analyzed the CO outcome in the absence of the Srs2 and Sgs1 helicases, DNA damage checkpoint proteins as well as in a mutant proliferating cell nuclear antigen (PCNA) and found that they all contribute to genome stability. Remarkably high effects on COs are mediated by srs2Delta, mrc1Delta and a pol30-RR mutation in PCNA. Our results support the view that Mrc1 plays a specific role in DNA replication, promoting the Srs2 recruitment to PCNA independently of checkpoint signaling. Srs2 would prevent formation of double Holliday junctions (dHJs) and thus CO formation. Sgs1 also negatively regulates CO formation but through a different process that resolves dHJs to yield non-CO products.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
