

Novel transcripts of Nox1 are regulated by alternative promoters and expressed under phenotypic modulation of vascular smooth muscle cells
- PMID: 16724959
- PMCID: PMC1550311
- DOI: 10.1042/BJ20060300
Novel transcripts of Nox1 are regulated by alternative promoters and expressed under phenotypic modulation of vascular smooth muscle cells
Abstract
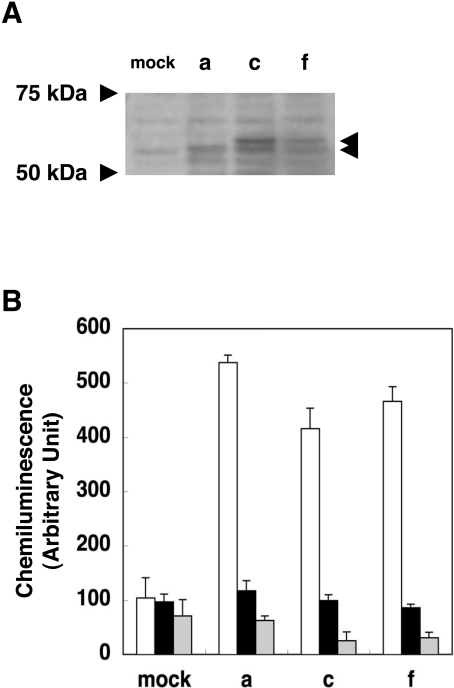
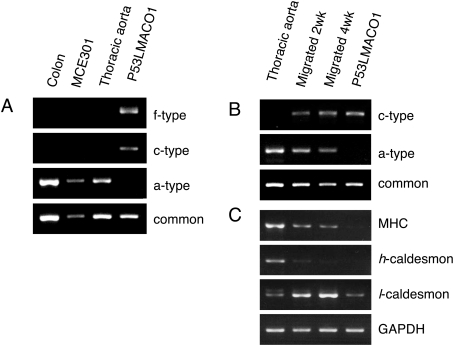
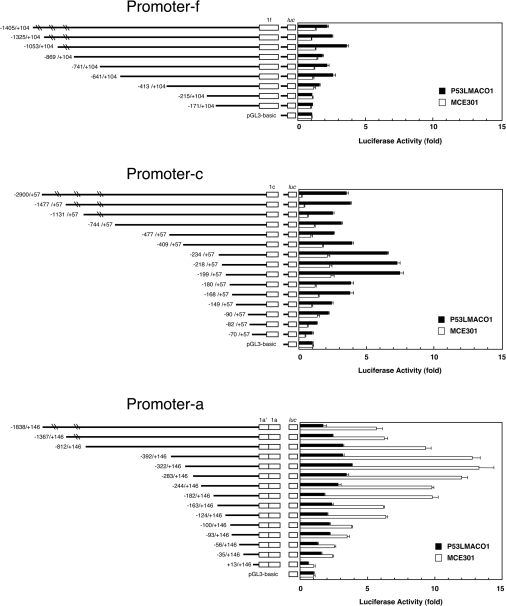
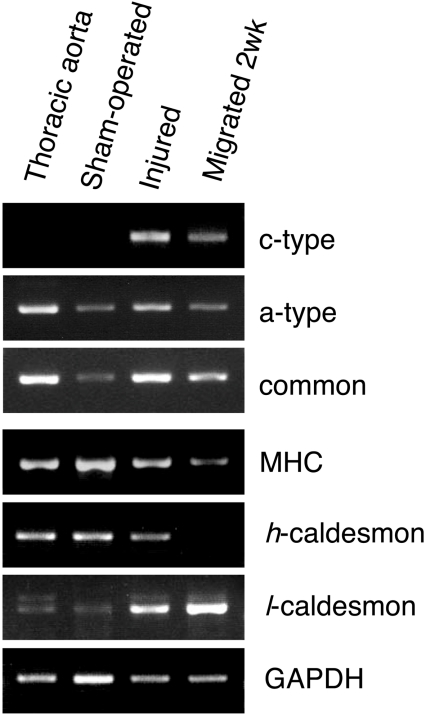
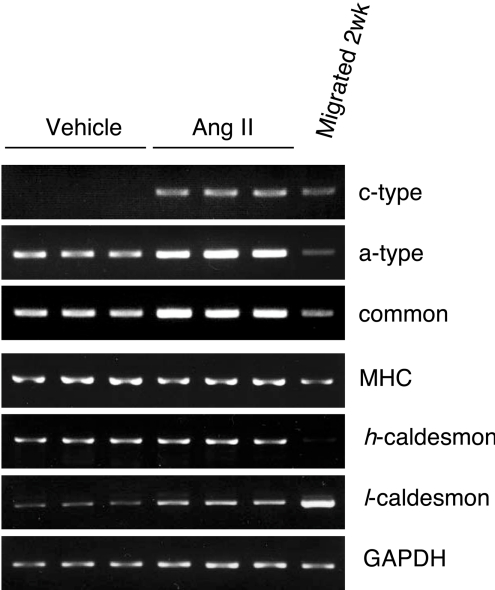
NADPH oxidase is implicated in the pathogenesis of various cardiovascular disorders. In vascular smooth muscle cells (VSMC), expression of NOX1 (NADPH oxidase 1), a catalytic subunit of NADPH oxidase, is low and is induced upon stimulation by vasoactive factors, while it is abundantly expressed in colon epithelial cells. To clarify the regulatory mechanisms underlying such cell-specific expression, the upstream regions directing transcription of the NOX1 gene were explored. In P53LMACO1 cells, a cell line originated from mouse VSMCs, two novel Nox1 mRNA species, the c- and f-type, were isolated. These transcripts contained 5'-untranslated regions that differed from the colon type mRNA (a-type) and encoded an additional N-terminal peptide of 28 amino acids. When these transcripts were fused to the c-myc tag and expressed in human embryonic kidney 293 cells, a fraction of translated proteins demonstrated the size containing the additional peptide. Proteins encoded by the c- and f-type mRNAs exhibited superoxide-producing activities equivalent to the activity of the a-type form. The a-type mRNA was expressed in the colon and in the intact aorta, whereas the c-type mRNA was detected in the primary cultured VSMCs migrated from aortic explants, in vascular tissue of a wire-injury model and in the thoracic aorta of mice infused with angiotensin II. The promoter region of the c-type mRNA exhibited transcriptional activity in P53LMACO1 cells, but not in MCE301 cells, a mouse colon epithelial cell line. These results suggest that expression of the Nox1 gene is regulated by alternative promoters and that the novel c-type transcript is induced under phenotypic modulation of VSMCs.
Figures

References
-
- Suh Y. A., Arnold R. S., Lassègue B., Shi J., Xu X., Sorescu D., Chung A. B., Griendling K. K., Lambeth J. D. Cell transformation by the superoxide-generating oxidase Mox1. Nature (London) 1999;401:79–82. - PubMed
-
- Bánfi B., Maturana A., Jaconi S., Arnaudeau S., Laforge T., Sinha B., Ligeti E., Demaurex N., Krause K. H. A mammalian H+ channel generated through alternative splicing of the NADPH oxidase homolog NOH-1. Science. 2000;287:138–142. - PubMed
-
- Lassègue B., Clempus R. E. Vascular NAD(P)H oxidases: specific features, expression, and regulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003;285:R277–R297. - PubMed
-
- Lassègue B., Sorescu D., Szöcs K., Yin Q., Akers M., Zhang Y., Grant S. L., Lambeth J. D., Griendling K. K. Novel gp91phox homologues in vascular smooth muscle cells: nox1 mediates angiotensin II-induced superoxide formation and redox-sensitive signaling pathways. Circ. Res. 2001;88:888–894. - PubMed
-
- Katsuyama M., Fan C., Yabe-Nishimura C. NADPH oxidase is involved in prostaglandin F2α-induced hypertrophy of vascular smooth muscle cells: induction of NOX1 by PGF2α. J. Biol. Chem. 2002;277:13438–13442. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases