

Characterization of E. coli tetrameric aldehyde dehydrogenases with atypical properties compared to other aldehyde dehydrogenases
- PMID: 16731973
- PMCID: PMC2242541
- DOI: 10.1110/ps.052039606
Characterization of E. coli tetrameric aldehyde dehydrogenases with atypical properties compared to other aldehyde dehydrogenases
Abstract
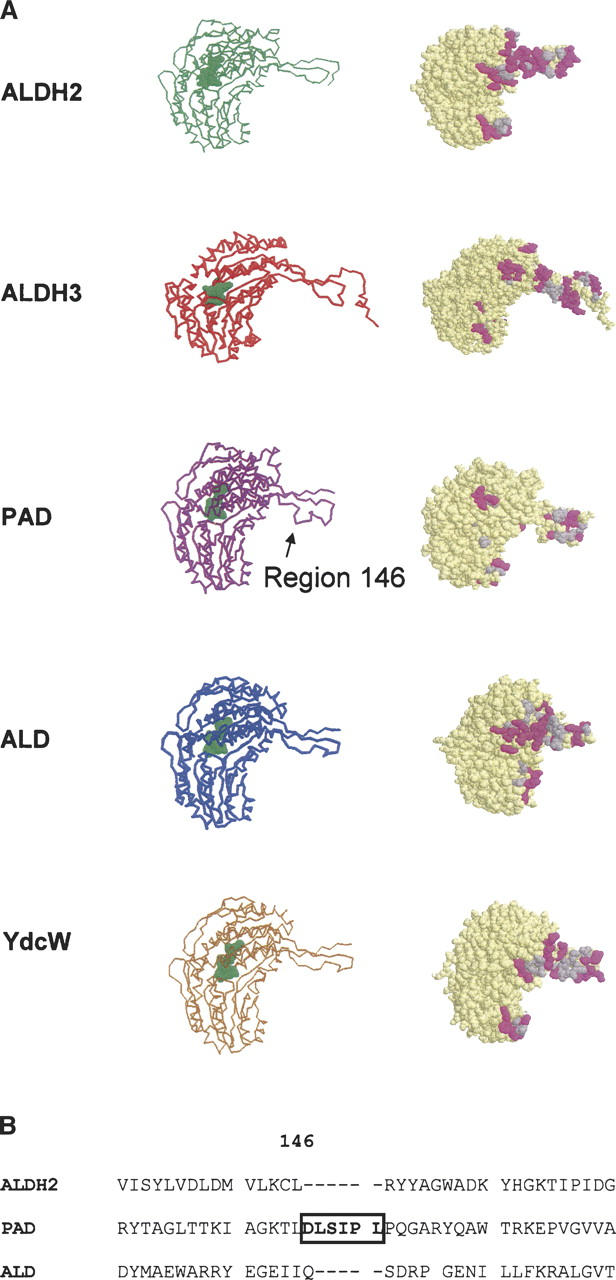
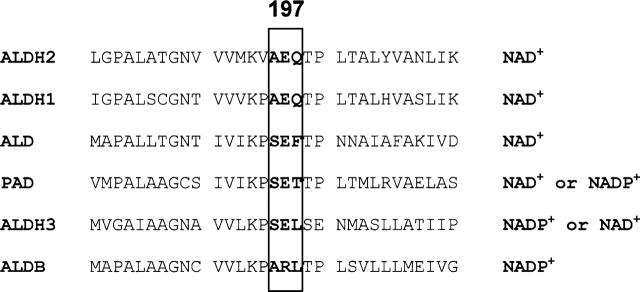
Aldehyde dehydrogenases are general detoxifying enzymes, but there are also isoenzymes that are involved in specific metabolic pathways in different organisms. Two of these enzymes are Escherichia coli lactaldehyde (ALD) and phenylacetaldehyde dehydrogenases (PAD), which participate in the metabolism of fucose and phenylalanine, respectively. These isozymes share some properties with the better characterized mammalian enzymes but have kinetic properties that are unique. It was possible to thread the sequences into the known ones for the mammalian isozymes to better understand some structural differences. Both isozymes were homotetramers, but PAD used both NAD+ and NADP+ but with a clear preference for NAD, while ALD used only NAD+. The rate-limiting step for PAD was hydride transfer as indicated by the primary isotopic effect and the absence of a pre-steady-state burst, something not previously found for tetrameric enzymes from other organisms where the rate-limiting step is related to both deacylation and coenzyme dissociation. In contrast, ALD had a pre-steady-state burst indicating that the rate-limiting step was located after the NADH formation, but the rate-limiting step was a combination of deacylation and coenzyme dissociation. Both enzymes possessed esterase activity that was stimulated by NADH; NAD+ stimulated the esterase activity of PAD but not of ALD. Finding enzymes that structurally are similar to the well-characterized mammalian enzymes but have a different rate-limiting step might serve as models to allow us to determine what regulates the rate-limiting step.
Figures


References
-
- Bhat P.V., Labrecque J., Boutin J.M., Lacroix A., Yoshida A. 1995. Cloning of a cDNA encoding rat aldehyde dehydrogenase with high activity for retinal oxidation Gene 166 303–306. - PubMed
-
- Brunner N., Siebers B., Hense L.R. 2001. Role of two different glyceraldehyde-3-phosphate dehydrogenases in controlling the reversible Embden-Meyerhof-Parnas pathway in Thermoproteus tenax. Regulation on protein and transcript level Extremophiles 5 101–109. - PubMed
-
- Caballero A., Baldoma L., Ros J., Boronat A., Aguilar J. 1983. Identification of lactaldehyde dehydrogenase and glycolaldehyde dehydrogenase as functions of the same protein in Escherichia coli J. Biol. Chem. 258 7788–7792. - PubMed
-
- Dickinson F.M. and Haywood G.W. 1986. The effects of Mg2+ on certain steps in the mechanisms of the dehydrogenase and esterase reactions catalysed by sheep liver aldehyde dehydrogenase. Support for the view that dehydrogenase and esterase activities occur at the same site on the enzyme Biochem. J. 233 877–883. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
