

The human Vps29 retromer component is a metallo-phosphoesterase for a cation-independent mannose 6-phosphate receptor substrate peptide
- PMID: 16737443
- PMCID: PMC1559457
- DOI: 10.1042/BJ20060033
The human Vps29 retromer component is a metallo-phosphoesterase for a cation-independent mannose 6-phosphate receptor substrate peptide
Abstract
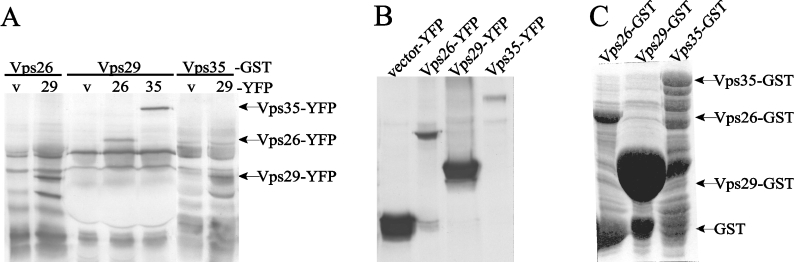
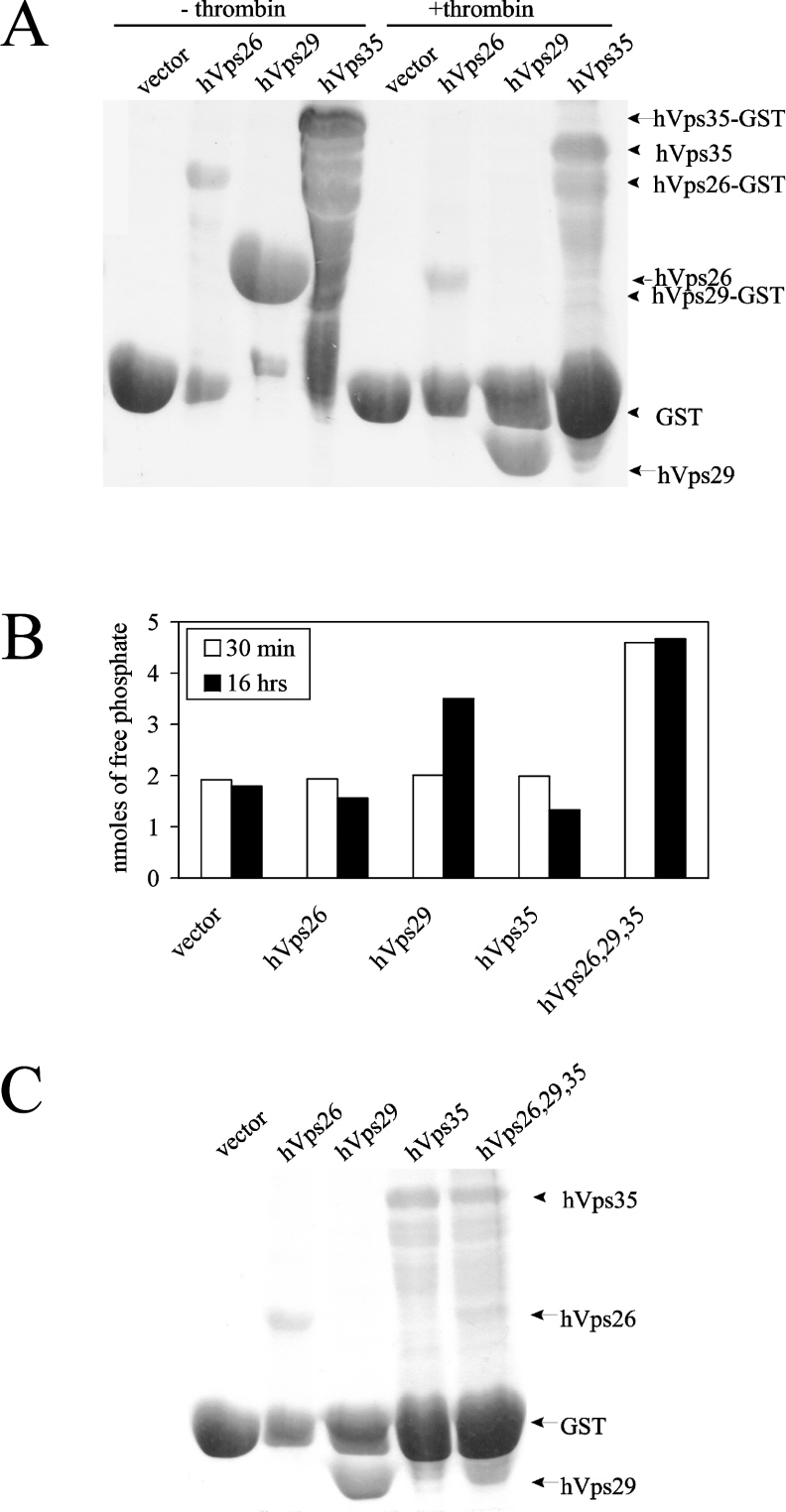
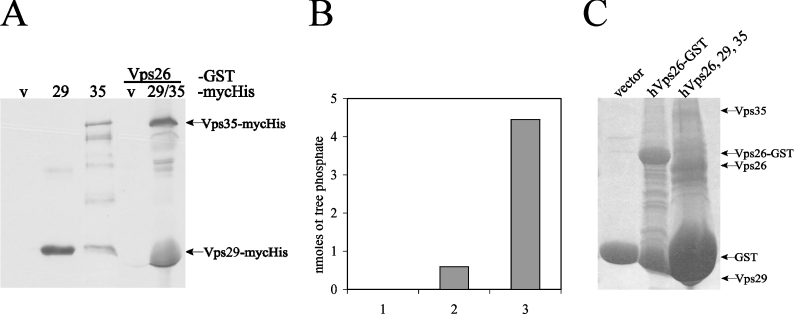
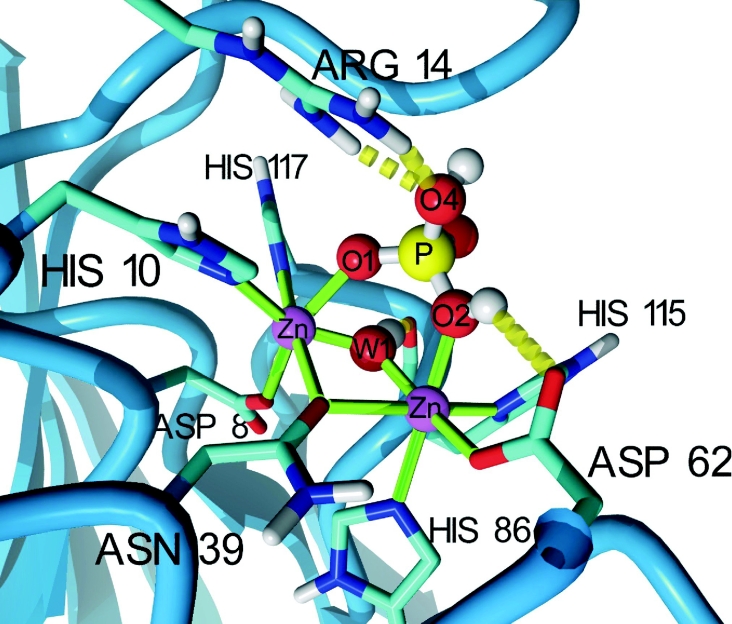
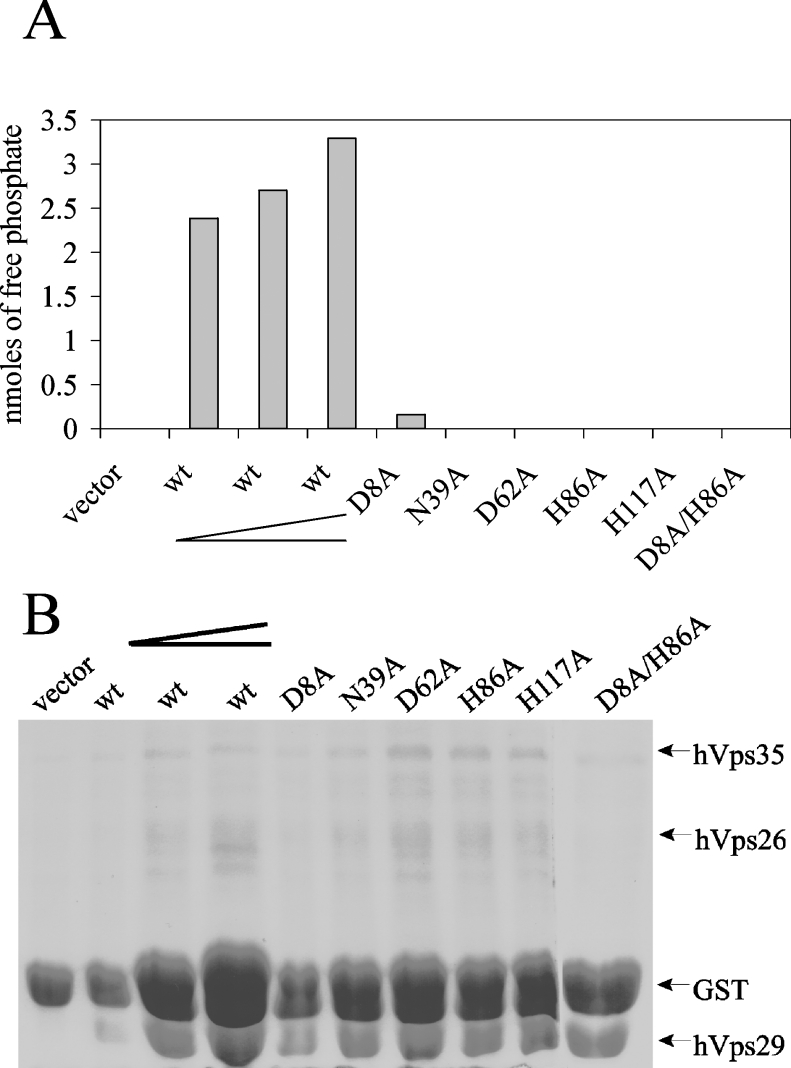
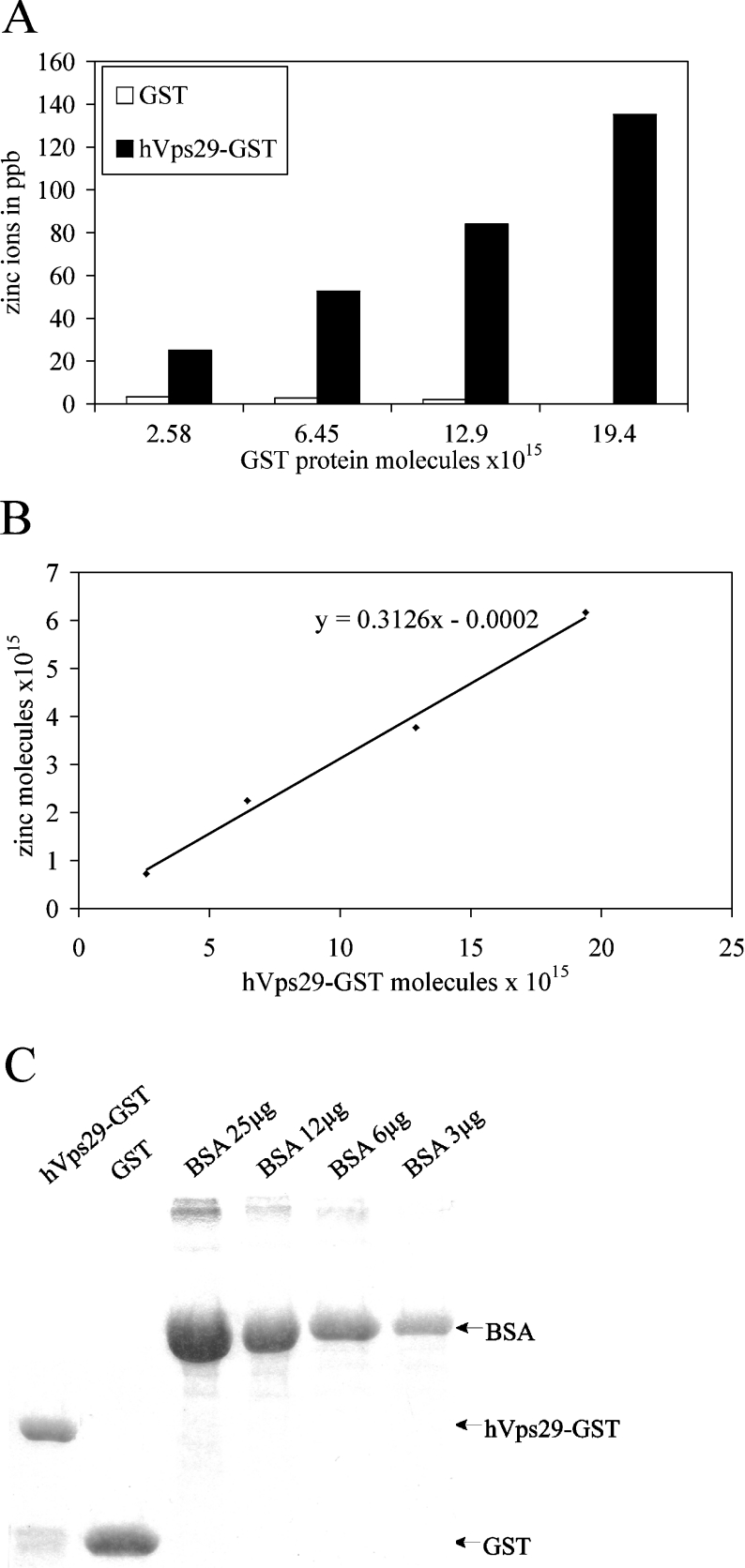
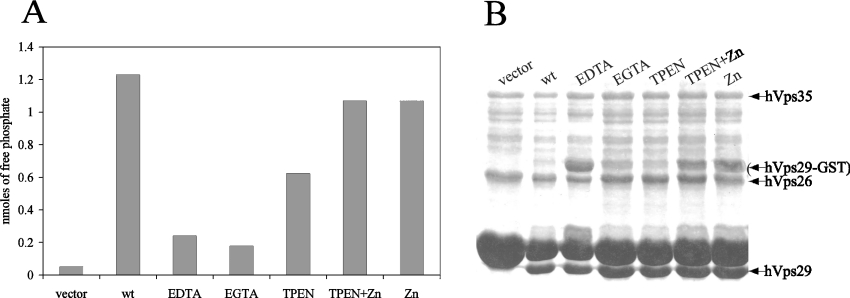
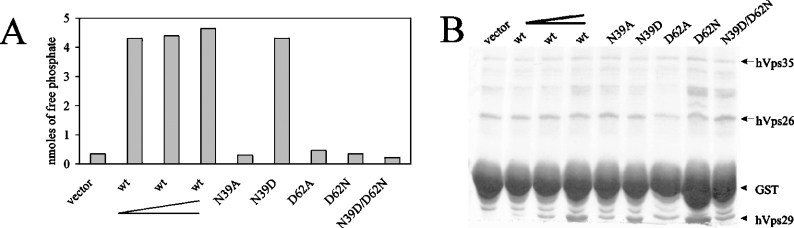
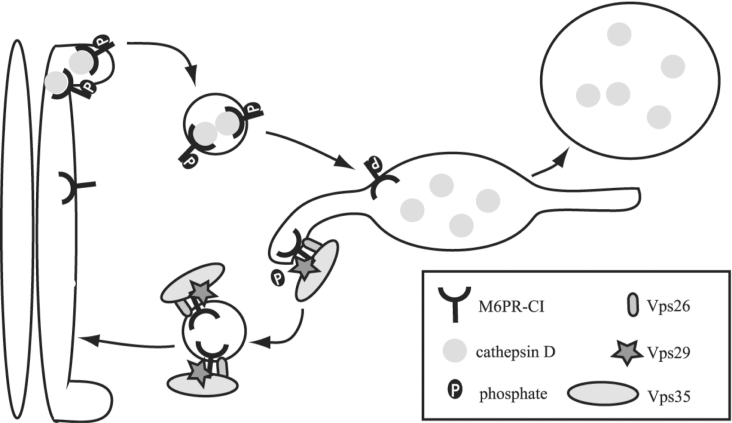
The retromer complex is involved in the retrograde transport of the CI-M6PR (cation-independent mannose 6-phosphate receptor) from endosomes to the Golgi. It is a hetero-trimeric complex composed of Vps26 (vacuolar sorting protein 26), Vps29 and Vps35 proteins, which are conserved in eukaryote evolution. Recently, elucidation of the crystal structure of Vps29 revealed that Vps29 contains a metallo-phosphoesterase fold [Wang, Guo, Liang, Fan, Zhu, Zang, Zhu, Li, Teng, Niu et al. (2005) J. Biol. Chem. 280, 22962-22967; Collins, Skinner, Watson, Seaman and Owen (2005) Nat. Struct. Mol. Biol. 12, 594-602]. We demonstrate that recombinant hVps29 (human Vps29) displays in vitro phosphatase activity towards a serine-phosphorylated peptide, containing the acidic-cluster dileucine motif of the cytoplasmatic tail of the CI-M6PR. Efficient dephosphorylation required the additional presence of recombinant hVps26 and hVps35 proteins, which interact with hVps29. Phosphatase activity of hVps29 was greatly decreased by alanine substitutions of active-site residues that are predicted to co-ordinate metal ions. Using inductively coupled plasma MS, we demonstrate that recombinant hVps29 binds zinc. Moreover, hVps29-dependent phosphatase activity is greatly reduced by non-specific and zinc-specific metal ion chelators, which can be completely restored by addition of excess ZnCl2. The binuclear Zn2+ centre and phosphate group were modelled into the hVps29 catalytic site and pKa calculations provided further insight into the molecular mechanisms of Vps29 phosphatase activity. We conclude that the retromer complex displays Vps29-dependent in vitro phosphatase activity towards a serinephosphorylated acidic-cluster dileucine motif that is involved in endosomal trafficking of the CI-M6PR. The potential significance of these findings with respect to regulation of transport of cycling trans-Golgi network proteins is discussed.
Figures
References
-
- Rothman J. H., Stevens T. H. Protein sorting in yeast: mutants defective in vacuole biogenesis mislocalize vacuolar proteins into the late secretory pathway. Cell. 1986;47:1041–1051. - PubMed
-
- Collins B. M., Skinner C. F., Watson P. J., Seaman M. N., Owen D. J. Vps29 has a phosphoesterase fold that acts as a protein interaction scaffold for retromer assembly. Nat. Struct. Mol. Biol. 2005;12:594–602. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous