

Polyadenylation of ribosomal RNA in human cells
- PMID: 16738135
- PMCID: PMC1474067
- DOI: 10.1093/nar/gkl357
Polyadenylation of ribosomal RNA in human cells
Abstract
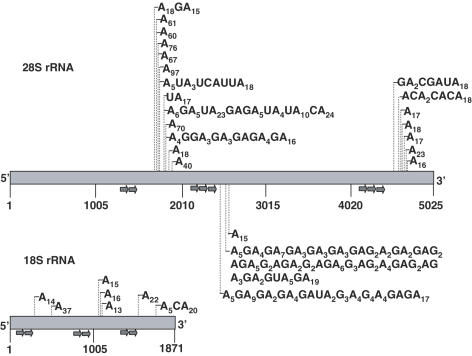
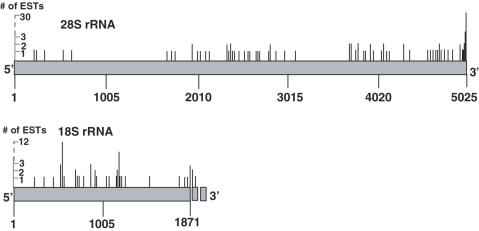
The addition of poly(A)-tails to RNA is a process common to almost all organisms. In eukaryotes, stable poly(A)-tails, important for mRNA stability and translation initiation, are added to the 3' ends of most nuclear-encoded mRNAs, but not to rRNAs. Contrarily, in prokaryotes and organelles, polyadenylation stimulates RNA degradation. Recently, polyadenylation of nuclear-encoded transcripts in yeast was reported to promote RNA degradation, demonstrating that polyadenylation can play a double-edged role for RNA of nuclear origin. Here we asked whether in human cells ribosomal RNA can undergo polyadenylation. Using both molecular and bioinformatic approaches, we detected non-abundant polyadenylated transcripts of the 18S and 28S rRNAs. Interestingly, many of the post-transcriptionally added tails were composed of heteropolymeric poly(A)-rich sequences containing the other nucleotides in addition to adenosine. These polyadenylated RNA fragments are most likely degradation intermediates, as primer extension (PE) analysis revealed the presence of distal fragmented molecules, some of which matched the polyadenylation sites of the proximal cleavage products revealed by oligo(dT) and circled RT-PCR. These results suggest the presence of a mechanism to degrade ribosomal RNAs in human cells, that possibly initiates with endonucleolytic cleavages and involves the addition of poly(A) or poly(A)-rich tails to truncated transcripts, similar to that which operates in prokaryotes and organelles.
Figures


References
-
- Dreyfus M., Regnier P. The poly(A) tail of mRNAs: bodyguard in eukaryotes, scavenger in bacteria. Cell. 2002;111:611–613. - PubMed
-
- Kushner S.R. mRNA decay in prokaryotes and eukaryotes: different approaches to a similar problem. IUBMB Life. 2004;56:585–594. - PubMed
-
- Slomovic S., Portnoy V., Liveanu V., Schuster G. RNA polyadenylation in prokaryotes and organelles; different tails tell different tales. Crit. Rev. Plant Sci. 2006;25:65–77.
-
- Edmonds M. A history of poly(A) sequences: from formation to factors to function. Prog. Nucleic Acid Res. Mol. Biol. 2002;71:285–389. - PubMed
-
- Shatkin A.J., Manley J.L. The ends of the affair: capping and polyadenylation. Nature Struct. Biol. 2000;7:838–842. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
