

Activation of muscarinic and nicotinic acetylcholine receptors in the nucleus accumbens core is necessary for the acquisition of drug reinforcement
- PMID: 16738243
- PMCID: PMC6675236
- DOI: 10.1523/JNEUROSCI.4494-05.2006
Activation of muscarinic and nicotinic acetylcholine receptors in the nucleus accumbens core is necessary for the acquisition of drug reinforcement
Abstract
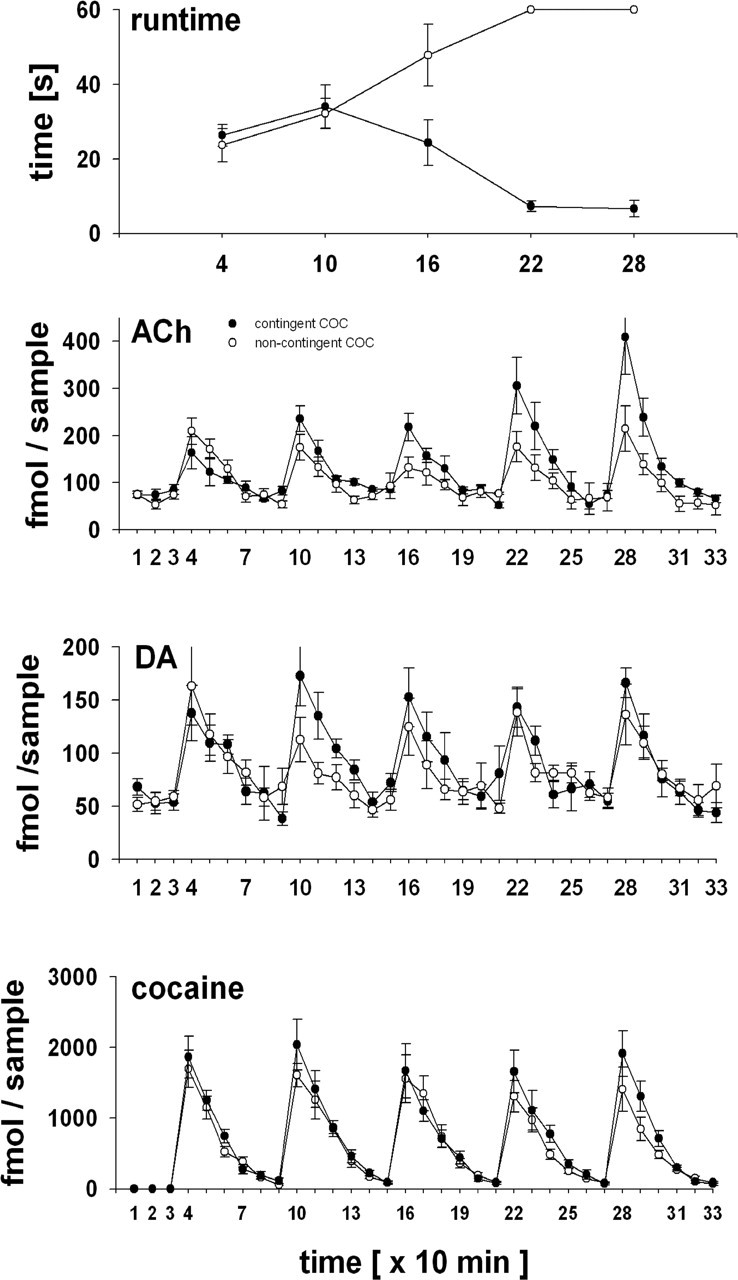
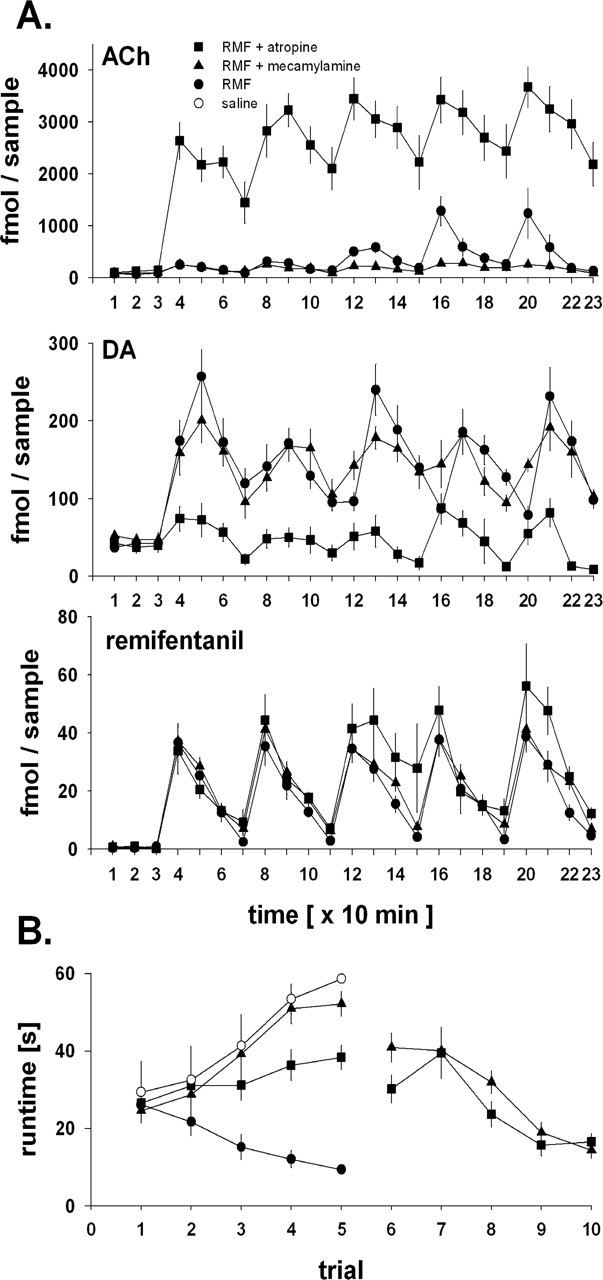
Neurotransmitter release in the nucleus accumbens core (NACore) during the acquisition of remifentanil or cocaine reinforcement was determined in an operant runway procedure by simultaneous tandem mass spectrometric analysis of dopamine, acetylcholine, and remifentanil or cocaine itself. Run times for remifentanil or cocaine continually decreased over the five consecutive runs of the experiment. Intra-NACore dopamine, acetylcholine, and drug peaked with each intravenous remifentanil or cocaine self-administration and decreased to pre-run baseline with half-lives of approximately 10 min. As expected, remifentanil or cocaine peaks did not vary between the five runs. Surprisingly, however, drug-contingent dopamine peaks also did not change over the five runs, whereas acetylcholine peaks did. Thus, the acquisition of drug reinforcement was paralleled by a continuous increase in acetylcholine overflow in the NACore, whereas the overflow of dopamine, the expected prime neurotransmitter candidate for conditioning in drug reinforcement, did not increase. Local intra-accumbens administration by reverse microdialysis of either atropine or mecamylamine completely and reversibly blocked the acquisition of remifentanil reinforcement. Our findings suggest that activation of muscarinic and nicotinic acetylcholine receptors in the NACore by acetylcholine volume transmission is necessary during the acquisition phase of drug reinforcement conditioning.
Figures

References
-
- Acquas E, Marrocu P, Pisanu A, Cadoni C, Zernig G, Saria A, DiChiara G (2001). Intravenous administration of 3,4-methylenedioxymethamphetamine (“Ecstasy”) enhances cortical and striatal acetylcholine in vivo. Eur J Pharmacol 418:207–211. - PubMed
-
- Bassareo V, DiChiara G (1999). Differential responsiveness of dopamine transmission to food-stimuli in nucleus accumbens shell/core compartments. Neuroscience 89:637–641. - PubMed
-
- Berridge KC, Robinson TE (2003). Parsing reward. Trends Neurosci 26:507–513. - PubMed
-
- Bhargava HN, Larsen AK, Rahmani NH, Villar VM (1993). Naltrexone-induced alterations of the distribution of morphine in brain regions and spinal cord of the rat. Brain Res 607:1–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials