

Leukemia-associated mutations within the NOTCH1 heterodimerization domain fall into at least two distinct mechanistic classes
- PMID: 16738328
- PMCID: PMC1489116
- DOI: 10.1128/MCB.01655-05
Leukemia-associated mutations within the NOTCH1 heterodimerization domain fall into at least two distinct mechanistic classes
Abstract
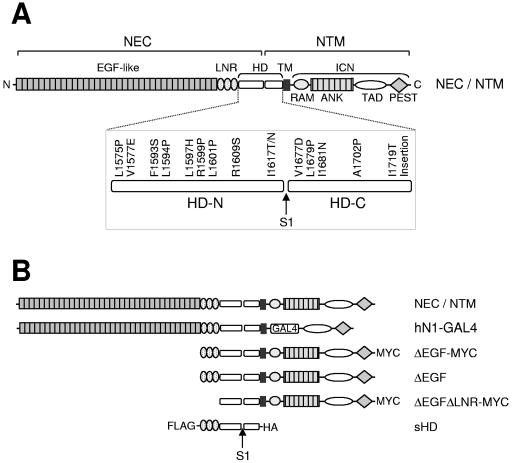
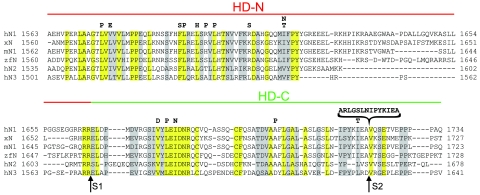
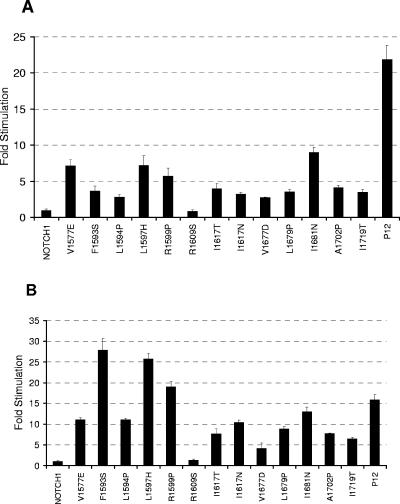
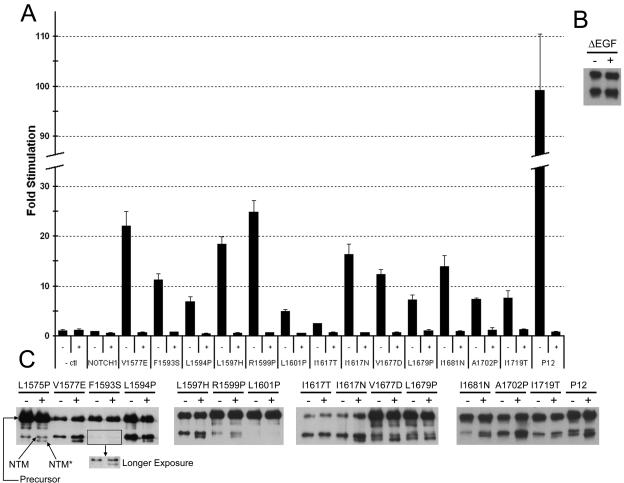
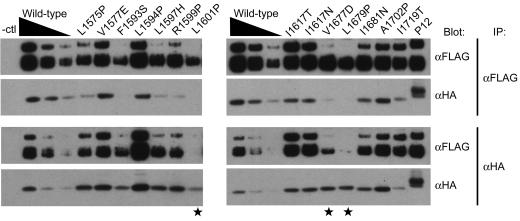
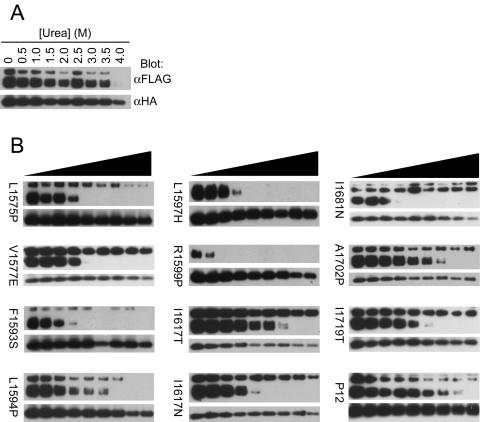
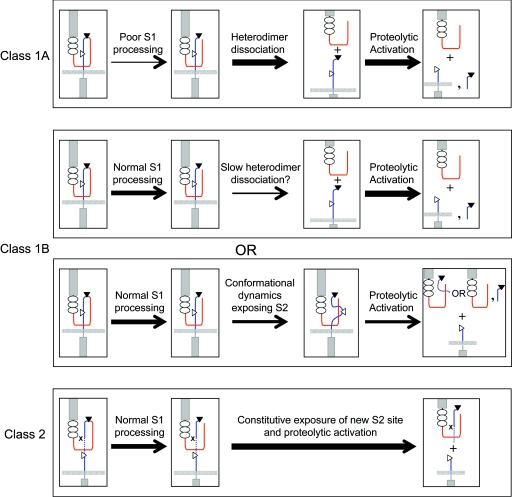
The NOTCH1 receptor is cleaved within its extracellular domain by furin during its maturation, yielding two subunits that are held together noncovalently by a juxtamembrane heterodimerization (HD) domain. Normal NOTCH1 signaling is initiated by the binding of ligand to the extracellular subunit, which renders the transmembrane subunit susceptible to two successive cleavages within and C terminal to the heterodimerization domain, catalyzed by metalloproteases and gamma-secretase, respectively. Because mutations in the heterodimerization domain of NOTCH1 occur frequently in human T-cell acute lymphoblastic leukemia (T-ALL), we assessed the effect of 16 putative tumor-associated mutations on Notch1 signaling and HD domain stability. We show here that 15 of the 16 mutations activate canonical NOTCH1 signaling. Increases in signaling occur in a ligand-independent fashion, require gamma-secretase activity, and correlate with an increased susceptibility to cleavage by metalloproteases. The activating mutations cause soluble NOTCH1 heterodimers to dissociate more readily, either under native conditions (n = 3) or in the presence of urea (n = 11). One mutation, an insertion of 14 residues immediately N terminal to the metalloprotease cleavage site, increases metalloprotease sensitivity more than all others, despite a negligible effect on heterodimer stability by comparison, suggesting that the insertion may expose the S2 site by repositioning it relative to protective NOTCH1 ectodomain residues. Together, these studies show that leukemia-associated HD domain mutations render NOTCH1 sensitive to ligand-independent proteolytic activation through two distinct mechanisms.
Figures
References
-
- Anderson, G., J. Pongracz, S. Parnell, and E. J. Jenkinson. 2001. Notch ligand-bearing thymic epithelial cells initiate and sustain Notch signaling in thymocytes independently of T cell receptor signaling. Eur. J. Immunol. 31:3349-3354. - PubMed
-
- Artavanis-Tsakonas, S., M. D. Rand, and R. J. Lake. 1999. Notch signaling: cell fate control and signal integration in development. Science 284:770-776. - PubMed
-
- Aster, J. C., E. S. Robertson, R. P. Hasserjian, J. R. Turner, E. Kieff, and J. Sklar. 1997. Oncogenic forms of NOTCH1 lacking either the primary binding site for RBP-Jkappa or nuclear localization sequences retain the ability to associate with RBP-Jkappa and activate transcription. J. Biol. Chem. 272:11336-11343. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases