

Post-transcriptional regulation of microRNA expression
- PMID: 16738409
- PMCID: PMC1484437
- DOI: 10.1261/rna.2322506
Post-transcriptional regulation of microRNA expression
Abstract
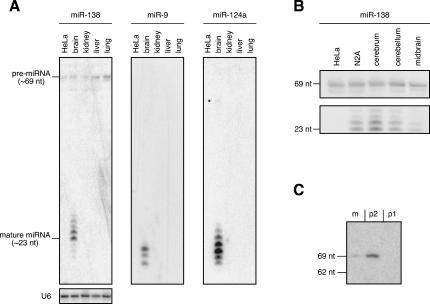
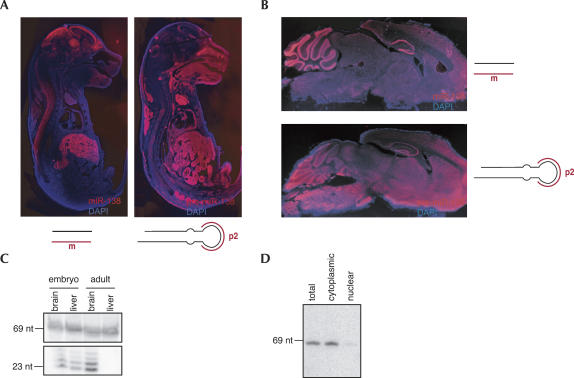
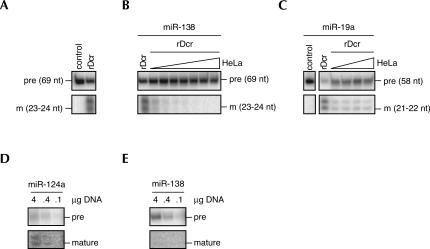
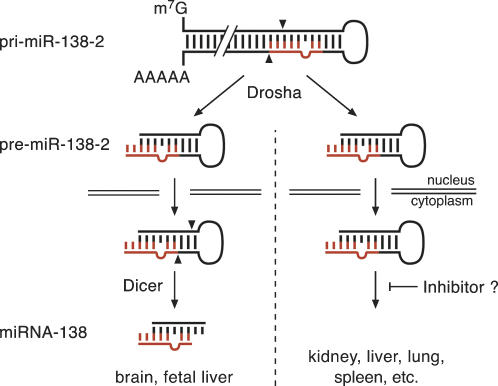
microRNAs (miRNAs) are endogenous, noncoding approximately 22-nucleotide RNA molecules that have recently emerged as fundamental, post-transcriptional regulators of cognate target gene expression. Many mammalian miRNAs are expressed in a tissue-specific manner, a phenomenon that has so far been attributed to transcriptional regulation. We here show by Northern blots and in situ hybridization experiments that the expression of mammalian miRNAs can be regulated at the post-transcriptional level. In particular, miR-138 is spatially restricted to distinct cell types, while its precursor, pre-miR-138-2, is ubiquitously expressed throughout all tissues analyzed. Furthermore, pre-miR-138-2 is exported from the nucleus to the cytoplasm, suggesting that cleavage of this pre-miRNA by Dicer is restricted to certain tissues and cell types. Thus, differential processing of pre-miRNAs might be an alternative mechanism to control miRNA function.
Figures
References
-
- Ambros V., Lee R.C., Lavanway A., Williams P.T., Jewell D. MicroRNAs and other tiny endogenous RNAs in C. elegans . Curr. Biol. 2003;13:807–818. - PubMed
-
- Bartel D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. - PubMed
-
- Bentwich I., Avniel A., Karov Y., Aharonov R., Gilad S., Barad O., Barzilai A., Einat P., Einav U., Meiri E., et al. Identification of hundreds of conserved and nonconserved human microRNAs. Nat. Genet. 2005;37:766–770. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources