

Noise resistance in the spindle assembly checkpoint
- PMID: 16738571
- PMCID: PMC1681502
- DOI: 10.1038/msb4100070
Noise resistance in the spindle assembly checkpoint
Abstract
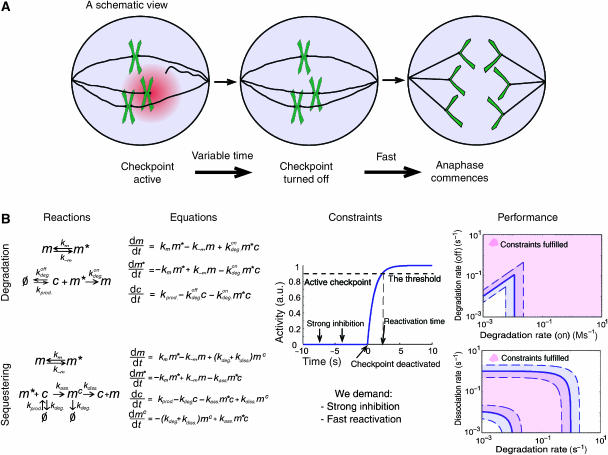
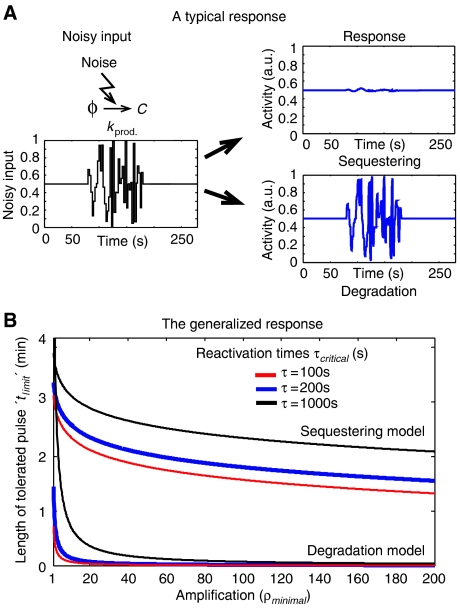
Genetically identical cells vary in the amount of expressed proteins even when growing under the same conditions. It is not yet clear how cellular information processing copes with such stochastic fluctuations in protein levels. Here we examine the capacity of the spindle assembly checkpoint to buffer temporal fluctuations in the expression of Cdc20, a critical checkpoint target whose activity is inhibited to prevent premature cell cycle progression. Using mathematical modeling, we demonstrate that the checkpoint can buffer significant fluctuations in Cdc20 production rate. Critical to this buffering capacity is the use of sequestering-based mechanism for inhibiting Cdc20, as apposed to inhibition by enhancing protein degradation. We propose that the design of biological networks is limited by the need to overcome noise in gene expression.
Figures
Similar articles
-
Phosphorylation of Cdc20 is required for its inhibition by the spindle checkpoint.Nat Cell Biol. 2003 Aug;5(8):748-53. doi: 10.1038/ncb1022. Nat Cell Biol. 2003. PMID: 12855955
-
Spindle checkpoint function requires Mad2-dependent Cdc20 binding to the Mad3 homology domain of BubR1.Exp Cell Res. 2006 Jun 10;312(10):1831-42. doi: 10.1016/j.yexcr.2006.02.018. Epub 2006 Apr 4. Exp Cell Res. 2006. PMID: 16600213
-
Cdc20 and Cks direct the spindle checkpoint-independent destruction of cyclin A.Mol Cell. 2008 May 9;30(3):290-302. doi: 10.1016/j.molcel.2008.02.027. Mol Cell. 2008. PMID: 18471975
-
Dynamics of spindle assembly and position checkpoints: Integrating molecular mechanisms with computational models.Comput Struct Biotechnol J. 2025 Jan 10;27:321-332. doi: 10.1016/j.csbj.2024.12.021. eCollection 2025. Comput Struct Biotechnol J. 2025. PMID: 39897055 Free PMC article. Review.
-
Noise in biology.Rep Prog Phys. 2014;77(2):026601. doi: 10.1088/0034-4885/77/2/026601. Epub 2014 Jan 20. Rep Prog Phys. 2014. PMID: 24444693 Free PMC article. Review.
Cited by
-
Noise propagation and signaling sensitivity in biological networks: a role for positive feedback.PLoS Comput Biol. 2008 Jan;4(1):e8. doi: 10.1371/journal.pcbi.0040008. Epub 2007 Dec 5. PLoS Comput Biol. 2008. PMID: 18179281 Free PMC article.
-
Homeorhesis and ecological succession quantified in synthetic microbial ecosystems.Proc Natl Acad Sci U S A. 2019 Jul 23;116(30):14852-14861. doi: 10.1073/pnas.1901055116. Epub 2019 Jul 10. Proc Natl Acad Sci U S A. 2019. PMID: 31292259 Free PMC article.
-
Mad3p, a pseudosubstrate inhibitor of APCCdc20 in the spindle assembly checkpoint.Genes Dev. 2007 Mar 15;21(6):655-67. doi: 10.1101/gad.1511107. Genes Dev. 2007. PMID: 17369399 Free PMC article.
-
Reverse engineering of the spindle assembly checkpoint.PLoS One. 2009 Aug 4;4(8):e6495. doi: 10.1371/journal.pone.0006495. PLoS One. 2009. PMID: 19652707 Free PMC article.
-
Types of potentials in a mitotic spindle.Biochem Biophys Rep. 2021 Jul 17;27:101076. doi: 10.1016/j.bbrep.2021.101076. eCollection 2021 Sep. Biochem Biophys Rep. 2021. PMID: 34337166 Free PMC article.
References
-
- Bar-Even A, Paulsson J, Maheshri N, O'Shea E, Pilpel Y, Barkai N (2006) Noise in protein expression scales with natural protein abundance. Nat Genet (in press) - PubMed
-
- Blake WJ, Kaern M, Cantor CR, Collins JJ (2003) Noise in eukaryotic gene expression. Nature 422: 633–637 - PubMed
-
- Brady DM, Hardwick KG (2000) Complex formation between Mad1p, Bub1p and Bub3p is crucial for spindle checkpoint function. Curr Biol 10: 675–678 - PubMed
-
- Cai L, Friedman N, Xie XS (2006) Stochastic protein expression in individual cells at the single molecule level. Nature 440: 358–362 - PubMed
-
- Chung E, Chen RH (2003) Phosphorylation of Cdc20 is required for its inhibition by the spindle checkpoint. Nat Cell Biol 5: 748–753 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
