

Could heat shock transcription factors function as hydrogen peroxide sensors in plants?
- PMID: 16740587
- PMCID: PMC2803459
- DOI: 10.1093/aob/mcl107
Could heat shock transcription factors function as hydrogen peroxide sensors in plants?
Abstract
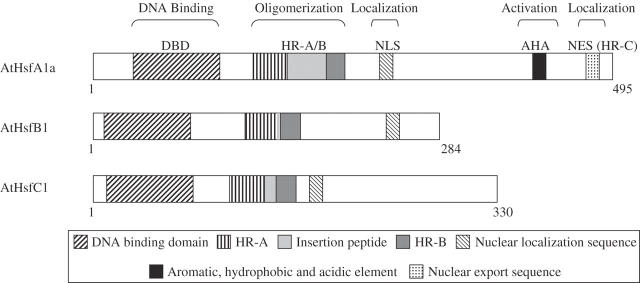
Background: Heat shock transcription factors (Hsfs) are modular transcription factors encoded by a large gene family in plants. They bind to the consensus sequence 'nGAAnnTCCn' found in the promoters of many defence genes, and are thought to function as a highly redundant and flexible gene network that controls the response of plants to different environmental stress conditions, including biotic and abiotic stresses. Hsf proteins encoded by different genes exhibit a high degree of complexity in their interactions. They can potentially bind and activate their own promoters, as well as the promoters of other members of their gene family, and they can form homo- or heterotrimers resulting in altered nuclear localization, as well as enhanced or suppressed transcription.
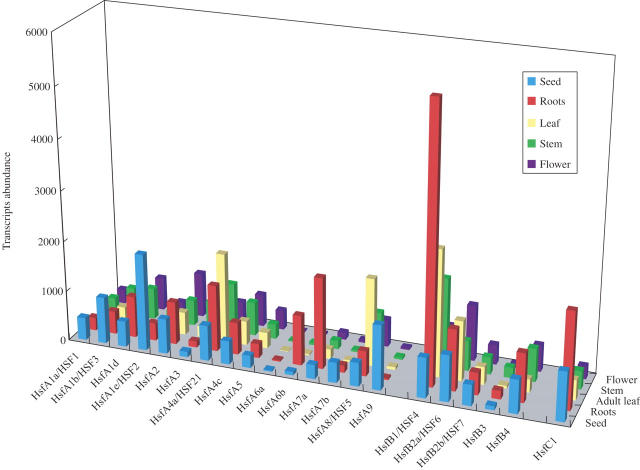
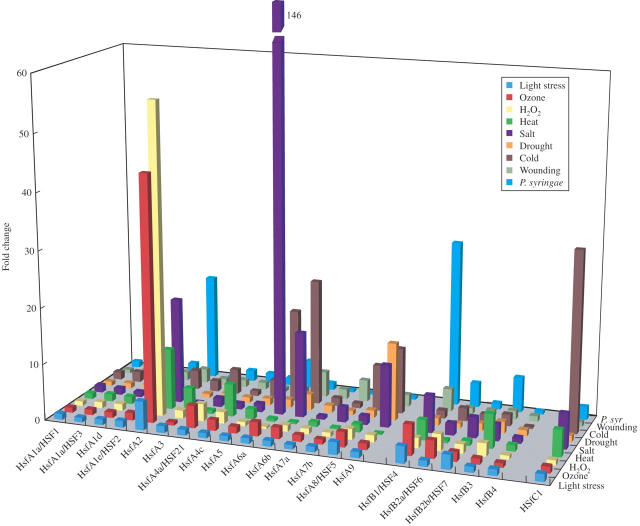
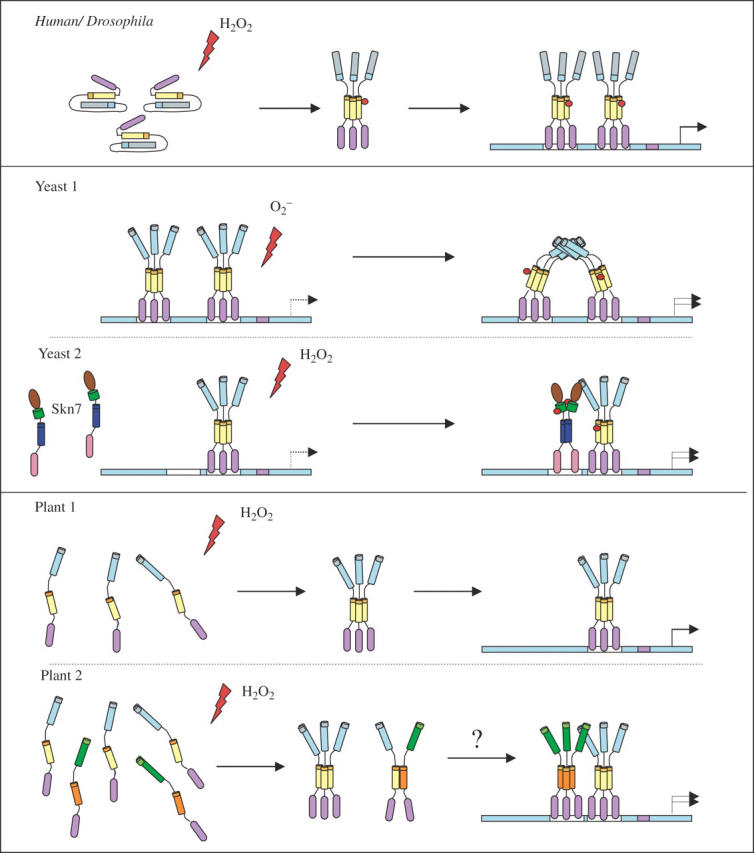
Scope: In this review, we summarize recent studies on Hsf function in Arabidopsis and tomato and present evidence obtained from microarray expression studies in Arabidopsis that the Hsf gene network is highly flexible and specialized, with specific members and/or member combinations controlling the response of plants to particular stress conditions. In addition, we describe recent studies that support the hypothesis that certain Hsfs function as molecular sensors that directly sense reactive oxygen species (ROS) and control the expression of oxidative stress response genes during oxidative stress.
Figures
References
-
- Apel K, Hirt H. 2004. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annual Review of Plant Biology 55: 373–399. - PubMed
-
- Baniwal SK, Bharti K, Chan KY, Fauth M, Ganguli A, Kotak S, et al.2004. Heat stress response in plants: a complex game with chaperones and more than twenty heat stress transcription factors. Journal of Biosciences 29: 471–487. - PubMed
-
- Bergmüller E, Porfirova E, Dörmann P. 2003. Characterization of an Arabidopsis mutant deficient in γ-tocopherol methyltransferase. Plant Molecular Biology 52: 1181–1190. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
